 2012, Vol. 48
2012, Vol. 48文章信息
- 任艳军, 马建军, 张立彬, 杜彬, 于凤鸣
- Ren Yanjun, Ma Jianjun, Zhang Libin, Du Bin, Yu Fengming
- 欧李叶表皮形态气孔指标与叶果矿质元素含量变化的关系
- Relationship between Leaf Epidermal Micro-Morphology and Stomata Indices and Mineral Elements Content Variations in Leaves and Fruits of Cerasus humilis
- 林业科学, 2012, 48(4): 133-137.
- Scientia Silvae Sinicae, 2012, 48(4): 133-137.
-
文章历史
- 收稿日期:2011-05-18
- 修回日期:2011-09-02
-
作者相关文章



欧李(Cerasus humilis)是我国特有的一种野生果树资源,其果实中钙、铁等元素尤为丰富(曹琴等,1999),同时具有抗旱、耐寒、耐贫瘠、耐盐碱等特性,是生态建设和防风固沙的良好树种(姜英淑等,2009; 李学强等,2009)。研究表明:叶片是植物进化过程中对环境变化敏感且可塑性较大的器官,叶表皮形态结构随着环境的变化而形成自身相对稳定的遗传特征,叶片的气孔、表皮毛、角质层等结构与植物的抗逆性密切相关(李芳兰等,2005; 李正理,1981; Bosabalidis et al., 2002; Sam et al., 2000; Mendes et al., 2001),而叶片气孔形态指标大小变化影响着植物的蒸腾、光合、呼吸等作用(潘瑞炽,2008)。马建军等(2010)研究发现:新梢不同部位叶片不同矿质元素含量变化存在明显差异,这可能与不同元素的生理特性,如移动性难易或转移再利用能力差异等因素有关。一般认为,主要运输动力之一蒸腾作用在影响和促进矿质元素在各组织器官中的吸收和分配发挥着重要作用,而叶片蒸腾作用能力的大小与叶片组织微形态结构如气孔大小和气孔密度等因素有关。因此,研究揭示欧李叶表皮形态及气孔特征在影响矿质元素代谢水平方面的作用具有重要意义。本文以同一生境条件下欧李为试材,利用扫描电镜方法观测欧李叶表皮细胞形态结构和气孔的形态指标大小,并探讨其与欧李叶果中矿质元素含量变化的关系,以期为揭示欧李矿质营养吸收转运机制提供基础理论依据。
1 材料与方法 1.1 试验材料欧李来源于燕山山脉野生欧李群体,实生繁殖并定植于河北科技师范学院园艺园林试验站野生欧李资源圃内,自然生长,始花期为4月中下旬,果实成熟期为7月下旬至8月上旬,株龄8年。
1.2 试验方法随机选取标记的28株表型性状差异明显(如在叶片和果实大小、形状及颜色等表型性状)的欧李株系作为研究材料,于7月中旬10:00(晴天)采摘不同株系不同方位结果基生枝中上部成熟健康叶片30片,装袋后带回实验室,随机选取5片叶片先用蒸馏水冲洗,再用超声波超声清洗10 min,以除去叶表面的污物。在叶片近中脉处剪取1 cm×1 cm大小叶块,用FAA固定液于4 ℃冰箱中固定24 h,再用乙醇进行梯度脱水,每次10 min,丙酮置换后,用临界点干燥仪(K850型)进行临界点干燥,离子溅射仪(SBC-12型)喷金处理2 min。利用扫描电子显微镜(KYKY-2800型)对欧李叶片上下表面组织形态特征进行观测,并利用扫描电镜软件对每个样品随机测量10个气孔的长度和宽度,计数单位面积内气孔个数以计算气孔密度,并计算其平均值。同时于果实成熟期采集与叶片样品相对应株系的果实样品,每株30个果实,按常规方法和要求将果实表面洗净,去果核后称取果肉并在65 ℃条件下烘干,计算单果含水百分率。烘干果肉与经70~80 ℃烘干的剩余叶片分别研磨处理后待测。采用HNO3-HClO4消解果实和叶片样品,分析测定Ca, Mg, K等矿质营养元素含量。
样品分析采用3200型原子吸收分光光度计测定,数据分析利用SPSS 17.00统计软件完成。
2 结果与分析 2.1 叶表皮微形态特征欧李群体叶片上、下表皮细胞的形态结构可各划分为2类。上表皮细胞,第一类是(类型Ⅰ)表皮细胞向下凹陷,大小不一,各细胞之间相互紧密连接形成蜂窝状(图 1A, B); 第二类是(类型Ⅱ)表皮细胞近圆形,向表面隆起(图 1C, D),同时均具有条纹状角质层; 上表皮均有表皮毛存在,表皮毛均为单细胞构成的单毛类型,表面光滑,基部较粗,末端渐细(图 1A, C),上表皮无气孔分布(图 1A, D); 不同株系之间表皮细胞向上隆起或向下凹陷的程度不同,表皮毛分布密度也存在差异。

|
图 1 野生欧李叶表皮结构扫描照片 Fig.1 The SEM pictures of leaf epidermal structure of wild C. humilis A, B:第一类型上表皮细胞结构; C, D:第二类型上表皮细胞结构; E:第一类型下表皮细胞结构; G:第二类型下表皮细胞结构; F, H:气孔形态 A, B: First type upper epidermal cell structure; C, D: Second type upper epidermal cell structure; E: First type lower epidermal cell structure; G: Second type lower epidermal cell tructure; F, H: Stomata characteristics |
下表皮细胞,一类是表皮细胞大小不等形状不规则,各细胞之间相互紧密连接形成沟壑状(图 1E); 另一类是表皮细胞为圆形,向表面隆起,细胞之间紧密连接(图 1G),下表面细胞向上隆起或向下凹陷的程度在不同株系之间也存在差异。下表皮无表皮毛存在,有大量气孔分布,气孔仅分布在叶脉之间,叶脉上方未见气孔,气孔突出于表皮细胞,呈椭圆形,无副卫细胞,保卫细胞由几个普通的表皮细胞不规则地围绕,属于无规则型,气孔外拱盖隆起呈脊状,表面光滑,内缘浅波状(图 1F, H)。同一株系叶片上下表皮细胞结构特征表现出一致性(类型Ⅰ或类型Ⅱ),可作为野生欧李遗传分类的指标之一。
由表 1可见:在实生类型群体中,平均气孔长度、宽度、气孔大小以及气孔密度分别为(8.17±1.37)μm,(2.60±0.68)μm, (21.99±9.17)μm2和(867.43±198.65) mm-2,不同叶表皮细胞类型间气孔器指标大小略有差异,方差分析结果显示各类型间无显著差异(P>0.05),其中类型Ⅰ气孔小而密度略大,类型Ⅱ气孔大而密度略小。观测结果显示:欧李不同株系间气孔器各项指标变异程度较大,变异系数以气孔大小变异最大,其次为气孔宽度和气孔密度,气孔长度变异较小。在同一生境条件下自然生长的野生欧李其叶表皮微形态特征存在较丰富的遗传多样性。
|
|
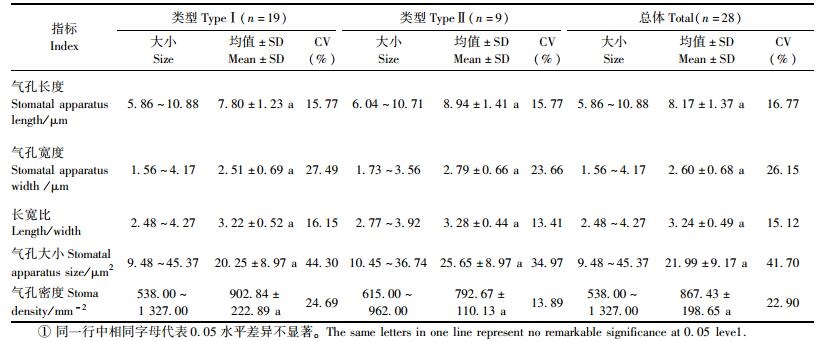
由表 2可见:在欧李实生群体中,矿质营养元素的含量变化,因组织器官、矿质元素种类以及株系不同其变异系数存在较大差异,叶片中元素含量变异系数的变幅为17.16%~78.20%,其中Na元素变异较大、Mn元素次之、Mg元素最小; 果实中元素含量变异系数的变幅为6.49%~103.30%,其中Mn元素变异较大、Fe元素次之、Na元素变异较小。叶片中除K元素显著低于果实外,其他元素含量均显著高于果实。
|
|

相关分析结果显示(表 3):欧李叶片气孔形态指标与叶片和成熟期果实中矿质营养元素的含量变化存在相关性,其相关程度大小因组织器官和元素种类不同而异。叶片中Ca元素含量与气孔宽度和气孔大小呈显著正相关关系,与气孔密度大小呈高度负相关(R= -0.414);叶片中Mg元素含量与气孔长度、气孔宽度以及气孔大小呈显著正相关关系; 叶片中其他元素含量与气孔器指标相关性均不显著。果实中Ca元素含量与气孔宽度呈显著正相关,与气孔大小呈高度正相关(R=0.402),与气孔长宽比呈极显著负相关; 果实中Zn元素含量与气孔宽度呈显著正相关,与气孔长宽比呈极显著负相关; 果实中Na元素含量与气孔密度呈显著正相关,果实中其他元素含量与气孔器指标的相关性均不明显。表明欧李叶片和果实中矿质营养元素含量水平差异与叶片气孔指标大小变化密切相关。
|
|
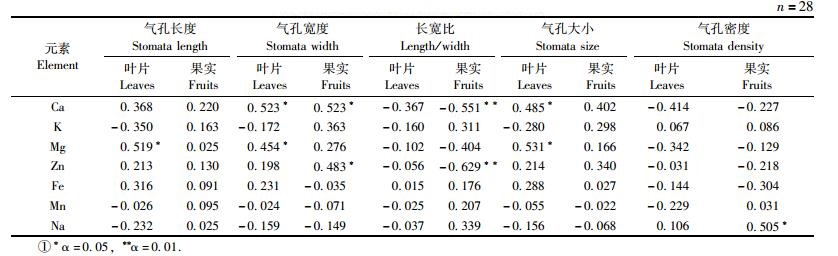
果树树体矿质元素运输主要是通过木质部导管内的交换机制来完成,木质部运输程度与元素梯度、运输形态、蒸腾拉力以及元素性质等因素有关(白昌华等,1989)。彭永宏(1991),钙素在椪柑(Citrus reticulata)树体中移动性很大,根部吸收的钙以上运为主,少有下运,运输主要受蒸腾作用控制并通过木质部实现的。已有的研究结果表明:欧李木质部、韧皮部和果实中水溶钙和果胶钙含量的年变化与相应器官中钙总量之间存在极显著正相关关系(马建军等,2008),且欧李果实中主要以水溶钙形态积累为主(马建军等,2007)。笔者研究认为:水溶钙形态是欧李树体中钙吸收和运输的主要钙形态,树体中水分的吸收和运输在影响钙离子吸收和运输方面发挥重要作用,而蒸腾作用是水分吸收和运输的主要动力。一般认为,成熟的叶片主要是通过气孔进行蒸腾,气孔蒸腾是植物蒸腾的主要方式,气孔宽而大,有利于气体的扩散,提高蒸腾速率,加快植物组织内水分的运转,从而促进欧李树体中水溶钙形态的吸收和运输,可能是造成果实发育期叶片中钙大量吸收和积累、新梢和果柄中水溶钙形态含量极显著增加(马建军等,2008),最后导致欧李成熟期果实中水溶钙形态的显著吸收积累的重要机制(马建军等,2007)。本研究相关分析结果表明:欧李叶片中的Ca,Mg元素含量和果实中的Ca,Zn,Na元素含量与叶表皮气孔指标大小变化存在显著或极显著的相关关系(表 3),其他元素相关性不显著,可能与不同种类的矿质元素在植物体内的转移再利用特性有关,需进一步深入研究。而叶片气孔密度与气孔长度(R=-0.560**)、气孔宽度(R=-0.629**)和气孔大小(R=-0.626**)均呈极显著负相关,表明欧李叶片气孔宽度和气孔大小变化是树体钙运输和果实钙吸收积累的重要影响因子。同时相关分析显示,欧李果实中含水百分率与叶片气孔长度(R=0.452*)、气孔宽度(R=0.559**)和气孔大小(R=0.533*)均呈显著或极显著的正相关关系,表明欧李果实含水百分率越高,果实中钙含量越高。由此推断,欧李叶片气孔形态指标大小差异是影响欧李树体矿质营养运输和吸收积累的重要影响因子,可能是欧李果实钙高效吸收的主要机制之一,该推论有待进一步研究证实。
| [] | 白昌华, 田华平. 1989. 苹果钙素营养研究进展综述. 西南农业学报, 2(3): 22–25. |
| [] | 曹琴, 杜俊杰, 刘和, 等. 1999. 野生欧李营养特性分析. 中国野生植物资源, 18(1): 34–36. |
| [] | 姜英淑, 陈书明, 王秋玉, 等. 2009. 干旱胁迫对2个欧李种源生理特征的影响. 林业科学, 45(6): 6–10. DOI:10.11707/j.1001-7488.20090602 |
| [] | 李芳兰, 包维楷. 2005. 植物叶片形态解剖结构对环境变化的响应与适应. 植物学通报, 22(增刊): 118–127. |
| [] | 李学强, 李秀珍. 2009. 盐碱胁迫对欧李叶片部分生理生化指标的影响. 西北植物学报, 29(11): 2288–2293. DOI:10.3321/j.issn:1000-4025.2009.11.021 |
| [] | 李正理. 1981. 旱生植物的形态和结构. 生物学通报(4): 9–12. |
| [] | 马建军, 于凤鸣, 张立彬, 等. 2010. 野生欧李果实发育期不同叶位叶片中主要矿质元素含量的变化. 果树学报, 27(1): 34–38. |
| [] | 马建军, 张立彬, 刘玉艳, 等. 2008. 野生欧李生长期组织器官中不同形态钙含量的变化及其相关性. 园艺学报, 35(5): 631–636. |
| [] | 马建军, 张立彬, 于凤鸣, 等. 2007. 野生欧李果实中不同形态钙的含量及分布. 园艺学报, 34(3): 755–759. |
| [] | 潘瑞炽. 2008. 植物生理学. 北京, 高等教育出版社: 16-24. |
| [] | 彭永宏. 1991. 应用45Ca研究椪柑对钙的吸收和运转. 西南农业大学学报, 13(5): 493–497. |
| [] | Bosabalidis A M, Kofidis G. 2002. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Science, 163(2): 375–379. DOI:10.1016/S0168-9452(02)00135-8 |
| [] | Sam O, Jeréz E, DellAmico J, et al. 2000. Water stress induced changes in anatomy of tomato leaf epidermis. Biologia Plantum, 43(2): 275–277. DOI:10.1023/A:1002716629802 |
| [] | Mendes M M, Gazarini L C, Rodrigues M L. 2001. Acclimation of Myrtus communis to contrasting Mediterranean light environments effects on structure and chemical composition of foliage and plant water relations. Environmental and Experimental Botany, 45(2): 165–178. DOI:10.1016/S0098-8472(01)00073-9 |