 2012, Vol. 48
2012, Vol. 48文章信息
- 李根, 周光益, 吴仲民, 梁瑞友
- Li Gen, Zhou Guangyi, Wu Zhongmin, Liang Ruiyou
- 南岭小坑木荷群落地上生物量
- Aboveground Biomass of a Naturaly-regenerated Schima superba Community at Xiaokeng of the Nanling Mountain
- 林业科学, 2012, 48(3): 143-147.
- Scientia Silvae Sinicae, 2012, 48(3): 143-147.
-
文章历史
- 收稿日期:2011-05-13
- 修回日期:2011-11-09
-
作者相关文章



2. 韶关市曲江区国营小坑林场 曲江 512162
2. Xiaokeng State Forestry Farm of Shaoguan Qujiang 512162
森林生物量是评价森林生态系统生产力、研究森林生态系统结构与功能的重要指标,也是深入了解森林生态系统变化规律的重要途径和评估森林碳收支的重要参数(吴仲民等,1998)。亚热带常绿阔叶林是我国面积最大的森林类型,它在世界森林植被中具有重要的作用和地位(张祝平等, 1996)。自20世纪90年代以来,很多学者研究了亚热带地区多种森林类型的生物量与生产力(张祝平等, 1989; 蚁伟民等, 2000; 莫江明等, 2004),南岭中亚热带常绿阔叶林物种丰富,特有种较多,素有“物种宝库”之称(祁承经等, 1992,但其生物量方面的研究较少。生物量的传统研究方法主要有皆伐法和相对生长法,仅见少数学者采用皆伐法估算海南热带林的生物量(黄全等, 1991; 李意德等, 1993),多数学者采用相对生长方程估算生物量,尤其是杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)、青冈(Cyclobalanopsis glauca)、檫木(Sassafras tsumu)等人工林(冯宗炜等, 1982; 冯宗炜, 1980; 柳江等, 2002)。本研究的目的就是利用800 m2皆伐样地的生物量数据,估算木荷天然次生林群落地上部分生物量,并建立主要优势种和混合树种的生物量相对生长模型,以期为开展南岭地区天然林碳汇研究并进一步揭示该森林类型的结构与功能提供基础数据,填补木荷群落天然林生物量研究方面的空白。
1 试验地概况研究地位于广东省韶关市曲江区东南部的国营小坑林场,地处南岭山地的大庾岭南缘(113°49′08″—113°52′12″ E,24°39′42″—24°42′33″N),以中山和丘陵为主。该地区属中亚热带季风气候区, 全年气候温和,年均气温20.3 ℃,年日照时数1 706 h,海拔200~1 301 m。全年无霜期约350天。年均降雨量1 530 mm,年均相对湿度79%。地带性土壤为山地红壤,成土母质为变质花岗岩。地带性植被类型为中亚热带常绿阔叶林(黄志宏等,2009)。现存植被为20~30年生天然次生林群落,优势树种为木荷(Schima superba)和杉木。本试验点位于小坑林场大芒坑林区,海拔约260 m,坡向西南,坡度为28°,调查研究的木荷群落是1989年采伐后自然更新的次生林。
2 研究方法样地调查:2009年9月,在小坑林场大芒坑林区选择1块800 m2的标准样地,将其分成8块10 m×10 m的样方,进行调查,对于胸径大于3 cm的树木,记录树种、胸径、树高、枝下高和冠幅,并计算乔木层各树种的立木密度、胸高断面积(BA)、平均胸径(D)、平均树高(H)和相对密度(RA)、相对频度(RF)、相对显著度(RD)以及重要值(IV)等指标。
生物量测定:2009年9月对800 m2的样地采用皆伐法获得生物量数据。在伐倒乔木之前,先进行下层植被生物量和凋落物测定;在每个10 m×10 m的样方内,均布设5个1 m2的小样方,收集并测定小样方内凋落物层生物量;下层植被生物量是将每个100 m2的样方内所有DBH < 3 cm的乔木幼树、幼苗、灌木和所有草本植物收集起来一起称量并取样。胸径≥3.0 cm的乔木按树木编号(1~210),依次伐倒,就地称取树干和枝叶的鲜质量;同时,对各植物器官进行取样称量,样品带回实验室在85 ℃恒温下烘至绝干,算出含水率,最后,算出所有树木各器官生物量干质量。
数据处理:用Excel 2010和SPSS 17.0进行数据计算及统计分析。
3 结果与分析 3.1 群落学特征分析木荷群落是杉木林皆伐后自然更新的森林群落,结构较为复杂,可分为乔木层(Ⅰ,Ⅱ)、灌木层、草本层以及层间植物,群落高度15~17 m,乔木层的主要优势树种为木荷、杉木,其次为微毛山矾(Symplocos wikstroemiifolia)、灰岩润楠(Machilus calcicola)、南酸枣(Choerospondias axillaris)。林下灌木稀少,主要是两广杨桐(Adinandra chinensis)(表 1)。草本植物稀疏,有少量芒萁(Dicranopteris linearis),层间植物多为鸡血藤(Millettia reticulata)。
|
|
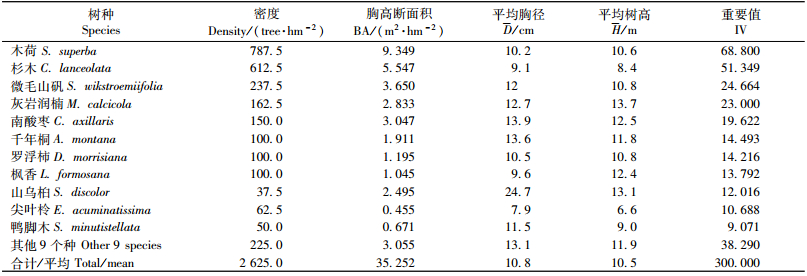
群落乔木层平均胸径10.8 cm,平均树高10.5 m,乔木树种20种,其中木荷、杉木、微毛山矾、灰岩润楠、南酸枣、千年桐(Aleurites montana)、罗浮柿(Diospyros morrisiana)、枫香(Liquidambar formosana)、山乌桕(Sapium discolor)、尖叶柃(Eurya acuminatissima)和鸭脚木(Schefflera minutistellata)11个树种的重要值之和为261.71,占乔木层的87.24%,另外9个树种占12.76%。
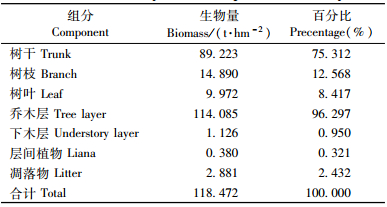
3.2 生物量组成特征1) 生物量在森林各层次中的分配根据皆伐实测生物量数据估算,该群落地上部分(AGB)总生物量为118.472 t·hm-2。乔木层地上生物量比重最大,达96.297%,下木层、层间植物和凋落物层分别占0.950%,0.321%和2.432%(表 2)。木荷-杉木群落乔木层地上部分总生物量为114.085 t·hm-2,其中树干生物量89.223 t·hm-2,树枝生物量14.890 t·hm-2,树叶生物量9.972 t·hm-2,各占乔木层地上部分生物量的78.2%,13.1%和8.7%。
|
|
2) 乔木层各树种地上部分生物量及其分配根据样地实测生物量数据(表 3),可知群落中木荷生物量所占比例最大,是乔木层地上部分生物量的33.3%;其他19个种的生物量占66.7%。群落中各树种地上部分生物量为0.329~38.031 t·hm-2;树干生物量0.252~28.996 t·hm-2,占乔木层地上部分生物量的72.2%~86.9%;树枝生物量0.043~5.081 t·hm-2,占乔木层地上部分生物量的6.7%~22.2%;树叶生物量0.019~3.955 t·hm-2,占乔木层地上部分生物量的5.2%~12.9%。
|
|

3) 乔木层各径阶地上部分生物量及其分配从图 1可以看出,群落乔木层地上部分生物量在各径阶及各器官中的分配规律,密度峰值在5~10 cm和10~15 cm径阶范围,生物量峰值在10~15 cm径阶范围;在10~20 cm径阶范围内的乔木层地上部分生物量占总乔木层地上部分生物量的71.8%,说明在此次生林中,地上部分生物量主要分布于10~20 cm径阶范围内。

|
图 1 木荷群落各径阶生物量及其分配 Fig.1 Distribution of tree biomass in DBH classes of S.superba community |
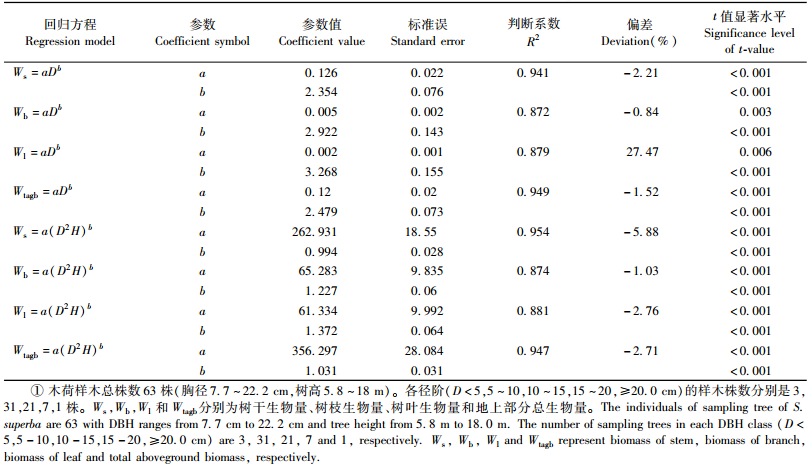
1) 木荷生物量回归模型分别以胸径(D)、胸径-树高(D2H)为变量,运用幂函数方程建立木荷各器官生物量回归模型(表 4)。可以看出,无论是以D还是以D2H为变量,树干和地上部分生物量回归模型的判断系数R2均高于树枝和树叶的R2;以D2H为变量的树干、树枝、树叶生物量回归模型的R2高于以D为变量的树干、树枝、树叶生物量回归模型的R2,说明D2H拟合的回归模型的精度要比D高。
|
|
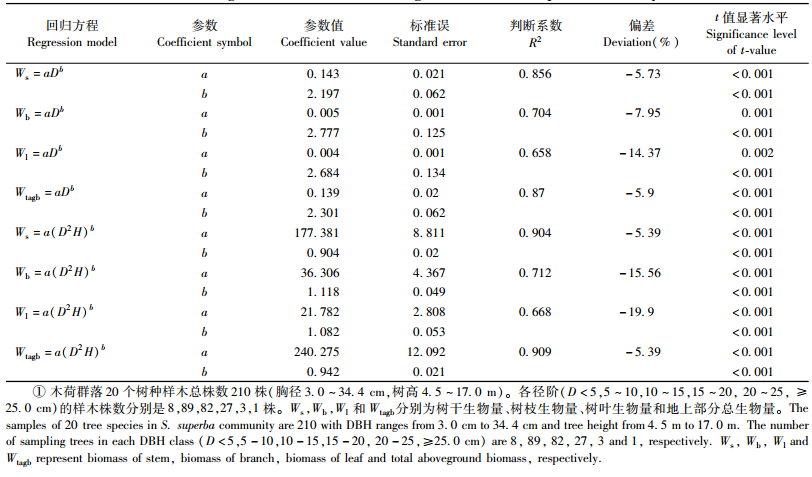
2) 混合种生物量回归模型分别以胸径(D)、胸径-树高(D2H)为变量,运用幂函数方程建立木荷群落各器官生物量回归模型(表 5)。可以看出,无论是以D还是以D2H为变量,树干和地上部分生物量回归模型的判断系数R2均高于树枝和树叶的R2,与实测值之间的偏差也远小于树枝和树叶,说明树干和地上部分总生物量比树枝和树叶生物量与变量D以及D2H的关系更加紧密;以D2H为变量树干、树枝、树叶和地上部分生物量回归模型的R2高于以D为变量树干、树枝、树叶生物量回归模型的R2,说明D2H拟合的回归模型的精度要比D拟合的高。
|
|
1) 本研究中的木荷群落是杉木林更新后的天然次生林群落,群落优势树种是木荷,生物量约占群落生物量的1/3;杉木密度最高,但处于被压状态,平均胸径和树高低于其他乔木,生物量也不足群落的1/10;除山乌桕外,其他树种的生物量与重要值成正比关系。
2) 该群落地上部分总生物量为118.472 t·hm-2,与广东鼎湖山的黄果厚壳桂(Cryptocarya concinna)群落(169.194 t·hm-2)、安徽肖坑的甜槠(Castanopsis eyrei)群落(120.165 t·hm-2)、浙江天童山的木荷群落(102.160 t·hm-2)地上生物量非常接近(张祝平等, 1996; 丁增发等, 2009; 杨同辉, 2005)。群落中乔木层比例最高,占96.3%,在乔木层各器官中树干生物量比例最高,占乔木层78.2%,枝叶生物量占乔木层的21.8%。因为林分郁闭度很高,该群落下木层、层间植物和凋落物层生物量非常低,为4.387 t·hm-2,只占群落生物量的3.7%。
3) 该群落密度峰值出现在5~15 cm径阶范围内,生物量峰值出现在10~20 cm径阶范围内,生物量峰值滞后于密度峰值,与周光益等(1997)研究结果一致。<5 cm和5~10 cm径阶枝叶生物量占地上生物量的13.6%和16.5%,>10 cm径阶枝叶生物量占地上生物量的22.3%~24.4%,说明<15 cm径阶乔木层枝叶生物量所占比重随胸径的增加而增加,>15 cm径阶乔木层枝叶生物量所占比重基本稳定,说明在树木生长过程中,枝叶生物量增长与树干生物量增长不同步。
4) 以D和D2H为变量的幂函数方程对树干和地上部分生物量的拟合度较优,这是因为影响树干生物量的主要因素是胸径、树高,且同它们之间基本呈正相关关系,而地上部分生物量中树干生物量比重非常高,占80%左右。影响枝叶生物量的因素很多,胸径、树高、林分密度、郁闭度、冠幅大小等对其都有较大影响,难以用胸径、树高拟合出相关关系很高的回归方程。综合考虑,以D2H为变量建立的回归模型拟合精度较D高,应用范围也更广泛;然而对于难以准确测定树高的林分,建议采用变量D拟合回归方程。
| [] | 丁增发, 姜春武, 汪小进, 等. 2009. 皖南山区北缘主要常绿林群落的生物量及生产力. 南京林业大学学报:自然科学版, 33(2): 129–133. |
| [] | 冯宗炜, 陈楚莹, 张家武, 等. 1982. 湖南会同地区马尾松林生物量的测定. 林业科学, 18(2): 127–134. |
| [] | 冯宗炜. 1980. 杉木人工林生物产量的研究.桃源综合考察报告集. 长沙, 湖南科学技术出版社: 322-333. |
| [] | 黄全, 李意德, 赖巨章, 等. 1991. 黎母山热带山地雨林生物量研究. 植物生态学与地植物学学报, 15(3): 197–206. |
| [] | 黄志宏, 田大伦, 周光益, 等. 2009. 广东南岭不同林分类型土壤养分状况比较分析. 东北林业大学学报, 37(9): 63–67. |
| [] | 李意德. 1993. 海南岛热带山地雨林林分生物量估测方法比较分析. 生态学报, 13(4): 313–320. |
| [] | 柳江, 洪伟, 吴承祯, 等. 2002. 天然更新的檫木林的生物量和生产力. 热带亚热带植物学报, 10(2): 105–120. |
| [] | 莫江明, 彭少麟, SandraBorwn, 等. 2004. 鼎湖山马尾松林群落生物量生产对人为干扰的响应. 生态学报, 24(2): 193–200. |
| [] | 吴仲民, 李意德, 曾庆波, 等. 1998. 尖峰岭热带山地雨林C素库及皆伐影响的初步研究. 应用生态学报, 9(4): 341–344. |
| [] | 蚁伟民, 张祝平, 丁明懋, 等. 2000. 鼎湖山格木群落的生物量和光能利用效率. 生态学报, 20(2): 397–403. |
| [] | 杨同辉. 2005. 浙江天童国家森林公园常绿阔叶林生物量研究. 东师范大学硕士学位论文. |
| [] | 张祝平, 丁明懋. 1996. 鼎湖山亚热带季风常绿阔叶林的生物量和光能利用效率. 生态学报, 16(5): 525–534. |
| [] | 张祝平, 彭少麟, 孙谷畴, 等. 1989. 鼎湖山森林群落植物量和第一性生产力的初步研究//中国科学院鼎湖山生态系统定位研究站. 热带亚热带森林生态系统研究. 第5集. 北京: 科学出版社, 63-73. |
| [] | 周光益, 曾庆波, 林明献, 等. 1997. 海南木莲人工林生物量及养分分配. 林业科学研究, 10(5): 453–457. |