 2012, Vol. 48
2012, Vol. 48文章信息
- 张明, 宋晓斌, 张星耀, 王振朝, 梁军
- Zhang Ming, Song Xiaobin, Zhang Xingyao, Wang Zhenchao, Liang Jun
- 脉冲磁场对松材线虫生理生化及超微结构的影响
- Effects of Pulsed Magnetic Field on the Biochemical and Physiological Characters and Ultrastructure of Pine Wood Nematode
- 林业科学, 2012, 48(3): 136-142.
- Scientia Silvae Sinicae, 2012, 48(3): 136-142.
-
文章历史
- 收稿日期:2011-05-24
- 修回日期:2011-09-12
-
作者相关文章



2. 西北农林科技大学林学院 杨凌 712100;
3. 河北大学电信学院 保定 071002
2. College of Forestry, Northwest A&F University Yangling 712100;
3. College of Electronic and Information Engineering, Hebei University Baoding 071002
松材线虫病又称松树枯萎病,是由病原松材线虫(Bursaphelenchus xylophilus)引起的一种毁灭性森林病害。随着国际贸易日益频繁,松材线虫随木包装以及木材的进出口,反复传播的可能性越来越大(张星耀等,2003),而传统的熏蒸、热处理和杀线虫剂处理效率低、易造成环境污染(陈劲松等,2010; 刘涛等,2010)。因此,经济、高效、环保的脉冲磁场技术在松材线虫病的防控可能性的研究尤为重要(刘亚宁,2002)。已有研究表明:脉冲磁场穿透能力极强,能全方位地作用于生物体,可直接影响到细胞的酶活性、膜的通透性,造成染色体畸变、细胞的结构和功能变化(刘亚宁,2002),使生物体内发生各种生理、生化和功能的变化(王保义等,1997),从而在一定条件下促进或抑制生物体的生长发育,甚至导致生物的死亡。
本文以松材线虫为研究对象,研究脉冲磁场对松材线虫活力、繁殖、酶活性以及体表特征等的影响,探寻脉冲磁场的生物效应机制,以及在实验室条件下有效抑制松材线虫生存的脉冲磁场技术参数,为其可行性研究及应用提供基础数据。
1 材料与方法 1.1 试验材料松材线虫为中国林业科学研究院森林生态环境与保护研究保所保存的虫株。在25 ℃于长满灰葡萄孢(Botrytis cinerea)的PDA培养基上接种松材线虫并培养繁殖7天,然后用贝尔曼漏斗法分离,分离液在2 000 r·min-1离心5 min,除去上清液,浓缩收集线虫(王保义等,1997)。以超纯水为介质配成松材线虫液,显微镜下观察测算单位体积松材线虫液的线虫总量、雌雄比以及幼虫成虫比(重复计算10次得平均数),制备的线虫液为200条·mL-1,雌雄比3.36,幼虫成虫比2.8。
黑松(Pinus thunbergii)采自中国林业科学研究院,选择4年生健康枝,松枝直径1 cm左右。用超纯水清洗枝条表面,洗净后在超纯水中剪成4 cm长的小段,擦干备用(梁军等,2008)。
1.2 脉冲磁场生成装置试验用脉冲磁场装置的主要可控参数有脉冲前沿宽度、脉冲周期、脉冲个数。停止时间和脉冲串个数。这些参数的调控直接对磁场强度起决定作用。脉冲前沿宽度T1在0.1~10 ms范围内以0.1 ms为单位可调; 脉冲周期T2以1ms为单位可调(最大值为200 ms); 脉冲个数即一个脉冲串内的脉冲个数,以1为单位可调(最大值为100 s); 停止时间即2个脉冲串之间的间歇时间,以1 s为单位可调(最大值为100 s); 脉冲串个数以1为单位可调(最大值为100)。磁场强度峰值是通过脉冲前沿宽度T1计算出来的。
根据脉冲磁场发生装置,1 ms脉冲前沿宽度时磁场强度峰值为720 Gs; 5 ms脉冲前沿宽度时磁场强度峰值为3 180 Gs; 10 ms脉冲前沿宽度时磁场强度峰值为5 080 Gs。
1.3 试验方法1) 试验设计以脉冲前沿宽度和处理时间作为处理因子,采用完全随机区组设计试验,脉冲前沿宽度设1,5,10 ms 3个水平; 处理时间设20,40,60 min 3个水平;以无磁场处理为对照。各处理重复3次,每重复8个离心管(每管1.5 mL),每管注入1 mL松材线虫液(200条·mL-1,幼虫成虫比2.8,雌雄比3.36)。脉冲周期80 ms,脉冲串个数100个,脉冲个数100个,停止时间1 s。
处理完毕后,观察统计不同脉冲磁场的处理对松材线虫活力、繁殖的影响。随后根据脉冲磁场对松材线虫活力和繁殖影响的结果,选择脉冲前沿宽度10 ms,不同处理时间处理后的松材线虫,进行松材线虫纤维素酶活性、迁移扩散的测定以及对松材线虫体表特征的观察。
2) 指标测定 ① 松材线虫的校正死亡率脉冲磁场处理后,立即将松材线虫静置于体视显微镜下,观察松材线虫的恢复情况,记录最早恢复正常活动的松材线虫的虫龄、所需时间,所有松材线虫趋于正常所需时间,采用校正死亡率评价松材线虫的生存状况(梁军等,2008)。采用针刺法判定松材线虫的死活,热杀死形态及体长法判断虫龄(谈家金等,2003; 杨宝君,2003),计算公式如下:
死亡率(%)=处理死虫数/处理总虫数×100%,
校正死亡率(%)=[1-处理活虫数/
(处理总虫数×对照存活率) ]×100%,
对照存活率(%)=对照活虫数/
对照总虫数×100%。
② 松材线虫的校正繁殖抑制率松材线虫在25 ℃于长满灰葡萄孢的PDA培养基上培养繁殖7天后用贝尔曼漏斗法分离,2 000 r·min-1离心5 min,除去上清液,浓缩收集线虫,在显微镜下观测松材线虫数量。
若处理后分离线虫数≤接种数量,则校正繁殖抑制率I(%)=100%;若处理后分离线虫数>接种数量,则校正繁殖抑制率I(%)=[1-(处理后分离线虫数-接种数)/(对照线虫数-接种数量) ]×100%(梁军等,2008)。
③ 松材线虫纤维素酶活性测定分别取1 mL脉冲磁场处理过的松材线虫液,室温下放置48 h,期间不时摇动,置于4 ℃冰箱保存备用(马海宾等,2009)。吸取待测酶液200 μL于2 mL离心管中,分别以1%的羧甲基纤维素钠(CMC)、微晶纤维素(MC)和水杨素为底物[均用乙酸缓冲液(pH值4.6) 配制]测定内切β-1,4-葡聚糖酶、外切β-1,4-葡聚糖酶和β-葡萄糖苷酶的活性,加入底物400 μL,50 ℃水浴保温1 h,再加入3 mol·L-1NaOH溶液400 μL,DNS试剂800 μL,颠倒混匀,置沸水浴中8 min。设空白对照,每处理重复3次,取平均值(马海宾等,2009)。490 nm下测定吸光度,并计算还原糖的含量,在以上反应条件下,以每分钟每毫升酶液产生1 μg葡萄糖的量为一个酶活性单位(索凤梅等,2004)。
④ 松材线虫校正迁移抑制率脉冲磁场处理后将松材线虫液及时滴入松枝上端套紧的乳胶管(2 cm)内,管底部放有灭过菌的脱脂棉,乳胶管上端用脱脂棉塞上保湿。将松枝下端插到放有无菌水的50 mL离心管中,插入深度为1 cm(Oku et al., 1989)。每4 h时收集1次水液,连续收集108 h,显微镜下观测收集水样中松材线虫数量及虫龄。记录初次检出松材线虫的时间、检出松材线虫的虫龄比例; 计算校正迁移抑制率以评价脉冲磁场对松材线虫迁移扩散的影响。
校正迁移抑制率I(%)=[1-处理后线虫累计迁移量/对照线虫累计迁移量]×100%
⑤ 扫描电镜观察脉冲磁场对松材线虫体表特征的影响脉冲磁场处理后分别将对照和处理的松材线虫迅速转入2.5%戊二醛固定,然后用80%,90%,100%的乙醇脱水,脱水时尽量避免搅动,CO2临界点干燥,粘样,离子溅射法镀膜,扫描电镜下观察并照相(赵博光等,2000)。
2 结果与分析 2.1 脉冲磁场对松材线虫生存的影响以校正死亡率评价脉冲磁场对松材线虫生存的影响。由表 1可见:3,4龄幼虫对脉冲磁场具有较好的抗性,在脉冲磁场处理后可最早恢复正常的活动能力,而2龄幼虫和雄虫的抗性最弱; 不同处理水平的脉冲磁场对松材线虫的校正死亡率具有不同程度的影响,其中脉冲前沿宽度1 ms,不同处理时间的脉冲磁场对松材线虫的生存影响较小。结果表明:松材线虫受到脉冲磁场作用后,会出现短暂的活动迟缓、扭动受阻等紊乱行为,静置一定时间后,松材线虫的活动能力会逐渐恢复; 但随着处理时间的增加,松材线虫活动能力下降越快,虫体复原所需时间也越长; 同时脉冲前沿宽度越大、处理时间越长,松材线虫的校正死亡率越高。
|
|
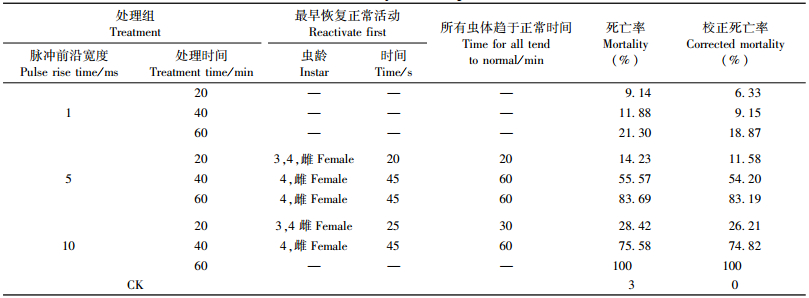
由图 1校正死亡率的均值分析,评价脉冲前沿宽度和处理时间对松材线虫生存影响的强弱。如图 1所示:脉冲前沿宽度1 ms对松材线虫的平均校正死亡率影响较小,但脉冲前沿宽度越大,松材线虫校正死亡率越高; 处理时间越长,松材线虫的校正死亡率也越高。即抑制松材线虫生存的最佳水平组合为脉冲前沿宽度10 ms、处理时间60 min。

|
图 1 不同脉冲前沿宽度和处理时间对松材线虫的平均校正死亡率的影响 Fig.1 The average corrected mortality of PWN treated by different pulse rise time and treatment time |
对松材线虫校正死亡率进行方差分析,脉冲前沿宽度和处理时间对松材线虫生存的影响均达到极显著水平。这说明脉冲前沿宽度和处理时间是抑制松材线虫生存的主要影响因子,两者的改变对松材线虫的生存有显著影响; 脉冲前沿宽度越大,处理时间越长,松材线虫的校正死亡率越高。对脉冲前沿宽度各水平的校正死亡率进行多重比较,结果表明:脉冲前沿宽度1与5 ms和脉冲前沿宽度1与10 ms处理的松材线虫的校正死亡率差异显著。对处理时间各水平的校正死亡率进行多重比较,结果表明:处理时间20与40 min和处理时间20与60 min处理的松材线虫的校正死亡率差异显著,处理时间40与60 min松材线虫的校正死亡率差异不显著。
2.2 脉冲磁场对松材线虫繁殖的影响适量的脉冲磁场处理可影响松材线虫的生长发育,抑制其繁殖。如表 2所示:脉冲磁场处理后由于松材线虫自身的恢复能力以及良好的繁育条件,脉冲前沿宽度1 ms时,不同处理时间的脉冲磁场对松材线虫的校正繁殖抑制率的影响极小,同样不同脉冲前沿宽度的脉冲磁场处理20 min对松材线虫繁殖的影响也较小。但随着处理时间的增加,松材线虫后期的繁育受到的抑制越明显,校正繁殖抑制率也越高。
|
|
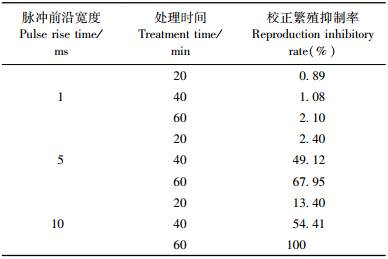
松材线虫平均校正繁殖抑制率的均值分析,以确定脉冲前沿宽度和处理时间各水平的优劣。如图 2所示:脉冲前沿宽度1 ms对松材线虫的平均校正繁殖抑制率几乎没有影响,但随着脉冲前沿宽度的增加,松材线虫的校正繁殖抑制率也越高; 同样处理时间越长,松材线虫的校正繁殖抑制率也越高。

|
图 2 脉冲前沿宽度和处理时间对松材线虫校正繁殖抑制率的影响 Fig.2 The reproduction inhibitory rate of PWN treated by different pulse rise time and treatment time |
对松材线虫校正繁殖抑制率进行方差分析,脉冲前沿宽度和处理时间对松材线虫繁殖的影响极显著,两者的改变对松材线虫的繁殖有显著影响。结果表明:脉冲前沿宽度和处理时间是抑制松材线虫繁殖的主要影响因子,脉冲前沿宽度越大,处理时间越长,松材线虫的校正繁殖抑制率越高。对脉冲前沿宽度各水平的校正繁殖抑制率进行多重比较,结果表明:脉冲前沿宽度1与5 ms和脉冲前沿宽度1与10 ms处理的松材线虫的校正繁殖抑制率差异显著。对处理时间各水平的校正繁殖抑制率进行多重比较,结果表明:处理时间20与40 min和处理时间20与60 min处理的松材线虫的校正繁殖抑制率差异显著,处理时间40与60 min处理的松材线虫的校正繁殖抑制率差异不显著。
2.3 脉冲磁场对松材线虫纤维素酶活性的影响侵入松树体内的松材线虫分泌的纤维素酶可分解细胞壁的纤维素,并能破坏木质部,在松树早期致病过程中起到了关键作用(田旺等,2005)。松材线虫分泌纤维素酶的3种组分包括:内切β-1,4-葡聚糖酶、外切β-1,4-葡聚糖酶和β-葡萄糖苷酶(马海宾等,2009)。如图 3所示:脉冲前沿宽度10 ms,不同处理时间的脉冲磁场处理后,与CK相比,内切β-1,4-葡聚糖酶、外切β-1,4-葡聚糖酶和β-葡萄糖苷酶的活性在脉冲磁场处理20 min后均出现上升,但随着处理时间的延长,纤维素酶活性逐渐降低,其中脉冲磁场处理60 min后纤维素酶活性降到最低值。

|
图 3 脉冲处理时间对松材线虫纤维素酶活性的影响 Fig.3 The effect of pulse rise time 10 ms and different treatment time on the cellulase activity of PWN |
如表 3所示:脉冲前沿宽度10 ms,不同处理时间的脉冲磁场处理后松材线虫的迁移扩散受到明显抑制,与对照相比,脉冲磁场处理后的松材线虫迁移出黑松枝条所需时间随处理时间的增加而延长,最长需88 h才可迁移出黑松枝条; 同时不同虫龄的松材线虫表现出不同的迁移现象,处理时间20和40 min均有3龄和4龄幼虫迁移出黑松枝条,由此可见3龄和4龄幼虫抗性较好,而脉冲磁场对2龄幼虫和雄虫迁移扩散的影响最为明显,无2龄幼虫和雄虫迁移出黑松枝条。
|
|
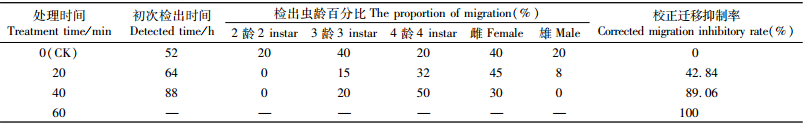
以校正迁移抑制率评价脉冲磁场对松材线虫迁移扩散的影响。图 4表明:在脉冲前沿宽度10 ms的条件下,随着脉冲磁场处理时间的增加,松材线虫的校正迁移抑制率越高,脉冲磁场对松材线虫迁移扩散抑制效果越好。

|
图 4 脉冲处理时间对松材线虫的校正迁移抑制率的影响 Fig.4 The effect of pulse rise time 10 ms and different treatment time on the corrected migration inhibitory rate of PWN |
扫描电镜观察,脉冲前沿宽度10 ms,不同处理时间的脉冲磁场处理后松材线虫的形态特征未发生明显变化,但其携带细菌的数量有明显变化。如图 5所示:松材线虫头部唇瓣的角质环包绕口孔,头部膨大,与体部连接处有明显的缢缩区; 雄虫尾部泄殖腔前后外缘稍向外突,具生殖乳突3对半; 雌虫尾部钝圆呈指状,无尾尖突。

|
图 5 脉冲磁场处理下松材线虫的形态特征 Fig.5 xSEM image of PWN by PMF 1,2,3. CK松材线虫CK of PWN; 4,5,6.脉冲磁场处理20 min后PWN by PMF treatment 20 min; 7,8,9.脉冲磁场处理40 min后PWN by PMF treatment 40 min; 10,11,12.脉冲磁场处理60 min后PWN by PMF treatment 60 min。图示圆圈表示松材线虫携带细菌的区域。The circle denotes the area that bacteria carried by PWN. |
CK松材线虫头部携带杆状细菌数量较多,尾部也携带有细菌(图 5-1~3)。脉冲磁场处理20 min后,松材线虫头部仍携带有杆状细菌,雄虫尾部肛前聚集不规则杂质,雌虫尾部也携带有个别细菌(图 5-4~6)。脉冲磁场处理40 min后,松材线虫头部、尾部未观察到细菌,雄虫尾部肛前有不规则杂质(图 5-7~9)。脉冲磁场处理60 min后,松材线虫头部、尾部清晰干净,未观察到细菌(图 5-10~12)。试验结果表明:脉冲磁场可抑制松材线虫体表携带细菌的生存,与CK相比脉冲磁场可导致松材线虫体表细菌数量的减少。
3 结论与讨论本研究表明:脉冲磁场可有效抑制松材线虫的生存活动能力、纤维素酶活性以及后期的繁殖。脉冲前沿宽度和处理时间是松材线虫校正死亡率和校正繁殖抑制率的主要影响因子,脉冲前沿宽度越大,处理时间越长,松材线虫的校正死亡率和校正繁殖抑制率越高。
短时间内松材线虫对脉冲磁场具有一定的抗性,出现短暂的紊乱行为,但随着脉冲前沿宽度的增加,脉冲处理时间的延长,虫体复原所需时间越长; 脉冲磁场可杀死松材线虫,其中脉冲前沿宽度10 ms,处理60 min的杀线效果最好,校正死亡率为100%。
脉冲磁场处理可抑制松材线虫在松枝内的迁移扩散,处理时间越长,对松材线虫迁移扩散的抑制效果越好。不同龄期的松材线虫的迁移扩繁能力不同,雌虫具有较好的抗性和迁移性,2龄虫和雄虫对脉冲磁场的抗性较差,未能迁移出松枝。松材线虫在脉冲磁场处理不同时间后,所分泌纤维素的酶活性出现短暂上升后逐渐降低,处理时间越长纤维素酶活性越低。
扫描电镜观察脉冲磁场对松材线虫体表特征的影响不明显,但对其携带杆状细菌的数量有着不同程度的影响。CK松材线虫的体表聚集较多的杆状细菌,而脉冲磁场处理后,松材线虫体表的细菌数量有一定数量的减少,脉冲磁场可抑制松材线虫体表携带细菌的生存。
不同水平的脉冲磁场处理后,由于松材线虫通过自身一定的感官知觉和行为活动的改变,作出防御脉冲磁场的反应,表现为急性和慢性2种生物效应,从而使松材线虫表现出不同程度的生存活动能力、纤维素酶活性和繁殖能力,可能归咎于松材线虫本身的补偿性、耐受性的能力(Bessho et al., 1995)。
| [] | 陈劲松, 吕国荣, 李惠军, 等. 2010. 福建口岸进口散装美国木片熏蒸效果浅析. 植物检疫, 24(3): 62–64. |
| [] | 梁军, 王翔凤, 宁少华, 等. 2008. 高压脉冲电流对松材线虫活力和繁殖的影响. 林业科学, 44(9): 106–112. |
| [] | 刘涛, 张凡华, 王跃进, 等. 2010. 异硫氰酸甲酯和硫酰氟混用杀灭松材线虫的增效研究. 植物检疫, 24(3): 1–4. |
| [] | 刘亚宁. 2002. 电磁生物效应. 北京, 北京邮电大学出版社. |
| [] | 马海宾, 梁军, 吕全, 等. 2009. 松材线虫与拟松材线虫分泌的纤维素酶系研究. 林业科学研究, 22(3): 397–400. |
| [] | 索凤梅, 林长春, 王浩杰, 等. 2004. 松墨天牛纤维素酶的研究Ⅰ. 纤维素酶性质研究.林业科学研究, 17(5): 583–589. |
| [] | 谈家金, 冯志新, 杜永部. 2003. 几种物质对松材线虫的作用. 四川林业科技, 24(4): 32–34. |
| [] | 田旺, 张奇, 杨文博, 等. 2005. 抗松材线虫纤维素酶单链抗体库的构建及筛选. 生物工程学报, 21(6): 900–905. |
| [] | 王保义, 徐润民, 杨杰斌. 1997. 瞬态电磁场或脉冲电磁场生物效应的机理研究. 电子学报, 25(3): 37–40. |
| [] | 杨宝君. 2003. 松材线虫病. 北京, 中国林业出版社. |
| [] | 张星耀, 骆有庆. 2003. 中国森林重大生物灾害. 北京, 中国林业出版社. |
| [] | 赵博光, 郭道森, 高蓉. 2000. 松材线虫携带细菌部位的电镜观察. 南京林业大学学报, 24(4): 69–71. |
| [] | Bessho K, Yamada S, Kunitani T, et al. 1995. Biological responses in Caenorhabditis elegans to high magnetic fields. Experientia, 51(3): 284–288. DOI:10.1007/BF01931113 |
| [] | Oku H, Shiraishi T, Chikamatsu K. 1989. Active defense as a mechanism of resistance in pine against pine wilt disease. Ann Phytopath Soc Japan, 55(5): 603–608. DOI:10.3186/jjphytopath.55.603 |