 2012, Vol. 48
2012, Vol. 48文章信息
- 蔡鸿娇, 王宏毅, 傅建炜, 江兴荣, 刘金海
- Cai Hongjiao, Wang Hongyi, Fu Jianwei, Jiang Xingrong, Liu Jinhai
- 寄主植物对芒果壮铗普瘿蚊为害的生理效应
- Physiological Responses of Mango Plants to Infestation by Procontarinai robusta
- 林业科学, 2012, 48(3): 100-104.
- Scientia Silvae Sinicae, 2012, 48(3): 100-104.
-
文章历史
- 收稿日期:2010-08-09
- 修回日期:2010-10-28
-
作者相关文章



2. 厦门出入境检验检疫局技术中心 厦门 361026;
3. 福建省农业科学院植物保护研究所
2. Inspection and Quarantine Technical Center, Xiamen Entry-Exit Inspection and Quarantine Bureau Xiamen 361026;
3. Institute of Plant Protection, Fujian Academy of Agricultural Sciences Fuzhou 350013
芒果壮铗普瘿蚊(Procontarinia robusta)是厦门市芒果(Mangifera indica)行道树和果园的主要害虫,于2000年在厦门市岛内首次发现,2003年鉴定为一新种并命名;属双翅目(Diptera)瘿蚊科(Cecidomyiidae)普瘿蚊属(Procontarinai)(李军等,2003)。2002年厦门市集美区和同安区发现该虫,2003年扩散到厦门市海沧杏林一带,并暴发成灾。2004年在漳州龙岩等地发现该虫,2005年相继在龙海、南安、泉州等周边地区发现(林振基等,2006)。2006—2007年对福建与台湾芒果树树冠主要害虫的调查表明:芒果壮铗普瘿蚊的发生量最大(游泳等,2009)。据调查,该害虫为害程度逐年加重,每年暴发成灾,但其入侵来源及机制不详。
芒果壮铗普瘿蚊严重影响芒果树的生长发育。其成虫在芒果嫩叶表面产卵,幼虫在叶肉内生长发育,刺激受害部位形成虫瘿,严重的单个叶片可达到500个虫瘿,新梢叶片受害率达100%;受害芒果叶片畸形,叶片正面残留大量的黑色瘿壳,严重影响植物叶片光合作用和植株的营养生长,并招致真菌为害,引起叶枯,致使叶片提前衰老、脱落,造成绿化景观的极大破坏(王伟新等,2005)。
植物新陈代谢的几个重要指标包括可溶性糖、可溶性蛋白、硝态氮、叶绿素的含量以及植物保护酶系统的活力。可溶性糖和蛋白质是植物生命活动必须的物质。硝态氮是植物最重要的氮源,植物体内硝态氮含量可以反映土壤氮素供应情况,也能够反映出植物的氮素营养状况,同时硝态氮经过代谢还原后能合成氨基酸和蛋白质。叶绿素是植物光合作用的主要成分,主要包括叶绿素a、叶绿色b;类胡萝卜素也是光合作用的必要成分。
植物的保护酶系统主要包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等。植物正常代谢过程以及在各种环境胁迫下均能产生活性氧和自由基,活性氧和自由基的积累会引起细胞结构和功能的破坏;而由SOD、POD、CAT等组成的植物保护酶系统是植物体内高效清除自由基的系统。SOD是活性氧清除系统中第一个发挥作用的抗氧化酶;SOD歧化超氧阴离子自由基生成H2O2和O2,在保护细胞免受氧化损伤过程中具有十分重要的作用。POD是植物体内活性较高的一种酶,它与呼吸作用、光合作用及生长素的氧化等都有密切关系,在植物生长发育过程中其活性不断发生变化,因此POD活力可反映某一时期植物体内代谢水平的变化。CAT活性与植物的代谢强度及抗病、抗寒能力等有一定关系。本试验通过测定芒果壮普瘿不同程度为害下芒果叶片营养物质(可溶性糖、可溶性蛋白质、硝态氮)含量、光合作用物质叶绿素等以及植物保护酶活力的影响,对植物能否对害虫产生抗逆性或者具有耐害性进行了探讨。
1 材料与方法 1.1 采集样品2010年2月17日至3月17日在福建省厦门市集美大学周边地区随机选择芒果树30株,芒果树胸径30 cm,为行道树,属于开放式管理,未使用过任何药剂。
采用剪枝法对每样株随机剪取长度大约为50 cm的枝条5个(习金根等,2006;张飞萍等,2007),合计150个枝条。枝条上2010年新梢还未完全形成,所剪取叶片均由2009年秋梢长出。
1.2 为害等级划分选择叶面积约为40 cm2,根据叶片受害的不同程度(每叶片虫瘿个体数量)分为4个等级: 1~50、51~100、101~200、>200;以无虫瘿为害的为对照组(CK)。每个等级取30个叶片,清洗干净,去除叶脉,剪碎混匀待测。每个等级重复3次。
1.3 测定指标及方法测定可溶性蛋白质、可溶性糖、硝态氮、叶绿色含量,过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)的活力,具体操作参考李合生(2000) 的方法。
1.4 数据处理1) 蛋白质含量(mg·g-1)=
2) 可溶性糖含量(%)=[从标准曲线查得糖的量×提取液体积(mL)×稀释倍数]/[测定用样品液的体积(mL)×样品质量(g)×106]×100。
3) 叶绿体色素含量(mg·g-1)=色素的浓度(C)×提取液的体积×

|
式中: Ca,Cb分别为叶绿素a和b的浓度(mg·L-1);Cx·c为类胡萝卜素的总浓度;A663, A646, A470分别为叶绿素提取液在波长663, 646和470 nm下的光密度。
4) 硝态氮含量(μg·g-1)=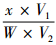
5) CAT活性采用高锰酸钾滴定法,CAT活性(mg·g-1min-1) =
6) POD活性(U·g-1min-1)=(ΔA470×VT)/(W×VS×0.01×t)。以每分钟内A470变化0.01为1个过氧化物酶活性单位(U)。式中: ΔA470为反应时间内光密度的变化;W为芒果叶片鲜质量,g;t为反应时间,min;VT为提取液总体积,mL;VS为测定时取用酶液体积,mL。
7) SOD总活性(U·mg-1)=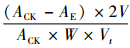

试验所得数据均以平均值±标准误表示,方差分析采用Duncan氏新复极差检验法,以DPS软件计算所得。
2 结果与分析 2.1 不同为害程度对寄主植物可溶性蛋白质、可溶性糖、硝态氮含量的影响根据试验结果,可溶性蛋白质含量标准曲线方程为y=0.013x-0.009(R2=0.999), 可溶性糖含量标准曲线方程为y=0.022x+0.011(R2=0.993), 硝态氮含量标准曲线方程为: y=0.075x+0.021(R2=0.996)。
从表 1可见:各为害等级可溶性蛋白质含量为0.65~0.80 mg·g-1, 显著低于对照组1.13 mg·g-1。可溶性糖含量和硝态氮含量在各等级间没有显著差异,可溶性糖含量为1.01%~1.31%,硝态氮含量为1 943.71~2 363.17 μg·g-1。
|
|

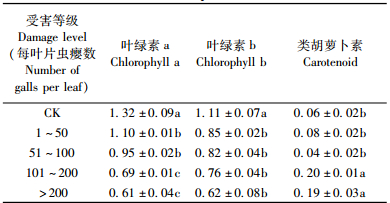
测定结果显示:受害后的叶片叶绿素a和叶绿素b含量均低于CK组,差异显著(表 2)。其中叶绿素a含量在为害等级为101~200与>200情况下显著低于其他等级,叶绿素b在各为害等级间差异不显著,均为0.62~0.85 mg·g-1。类胡萝卜素在等级为101~200与>200情况下显著高于其他为害等级,等级1~50与51~100情况下与CK没有显著差异(表 2)。
|
|
从表 3可见:CK组SOD,POD,CAT等保护酶系活性均显著低于各为害等级的活性。其中各为害等级叶片的SOD活性(251.87~412.94 U·g-1)显著高于CK组(157.13 U·g-1),而51~100与>200等级明显高于其他等级。
|
|

POD活性中,为害最严重的>200等级(1 331.25 U·g-1min-1)显著高于其他为害等级,CK组为27.08 U·g-1min-1,活力较低(表 3)。
CAT活性,>200等级(7.59 U·g-1min-1)显著高于其他等级。可以看出,随着为害程度的增加,CAT活性随之增加(表 3)。
3 讨论 3.1 不同为害程度对叶片营养物质含量的影响自然界中没有一种植物能够免受昆虫的取食为害,植物受害后,体内会发生各种新陈代谢的变化。植食性昆虫取食(Ohgushi, 2005)、人为对植物的损伤(Nykanen et al., 2004)、耕作方式的改变(Baumann et al., 2002) 都可诱导寄主植物营养水平的变化。植物可溶性糖类(主要是葡萄糖)积存于细胞中,其含量的增加对细胞的生命物质和生物膜起保护作用。植物组织中普遍存在的可溶性糖种类较多,常见的有葡萄糖、果糖、麦芽糖和蔗糖。一些研究表明害虫为害会引起植物可溶性糖含量降低,并引起次生代谢物质增加,如兴安落叶松鞘蛾(Coleophora obducta)取食为害落叶松(Larix sp.)林后,引起植物单宁增加(严善春等,2009),从而诱导植物产生局部防御系统,而单宁是很多植食性昆虫的拒食剂(王琪等,2008)。此外,人为损伤和松墨天牛(Monochamus alternatus)取食均能引起马尾松(Pinus massoniana)针叶内多糖和可溶性糖含量下降,其中虫伤叶中可溶性糖降低较多, 单宁和黄酮类物质含量会增加(李水清等,2007)。本研究表明受芒果壮铗普瘿蚊不同程度的为害,植物叶片内可溶性糖含量无显著差异,与前人的研究结果不一致,这可能由于,一方面所采集的叶片均为2009年老叶,所选择叶片的虫瘿密度基本形成与稳定后,叶片已基本完成发育,因此老叶新陈代谢减弱,可溶性糖含量基本保持稳定;另一方面也可能与害虫为害方式有关,瘿蚊为刺吸式为害,与鳞翅目蛾类幼虫、鞘翅目咀嚼式口器昆虫相比,对植物叶片的损伤较小。为害方式对植物体内可溶性糖含量的影响值得进一步探讨。
可溶性蛋白质是植物生命活动的指标,本研究表明随着叶片受害程度加大,可溶性蛋白质含量呈现逐渐下降的趋势;这与松墨天牛为害引起马尾松针叶可溶性蛋白质含量降低的报道一致(李水清等,2007)。植物体内可溶性蛋白质与其抗病性有关,植物被病菌侵染后,细胞内可溶性蛋白质含量会出现明显变化,如富贵竹(Dracalna sandeniansa)茎中可溶性蛋白含量与对细菌性茎腐病的抗病性呈正相关(李赤等,2007);另外,随着霜霉病(Plasmopara viticola)发病程度的逐渐加重,葡萄(Vitis vinifera)叶片内可溶性蛋白质含量也下降(齐慧霞等,2005)。因此,芒果树受该害虫为害后,导致其体内可溶性蛋白含量下降是必然的。
叶绿素含量是植物光合作用的主要指标,高等植物叶绿体中的叶绿素主要有叶绿素a和叶绿素b 2种。叶绿体中除含有叶绿素外也含有类胡萝卜素,类胡萝卜素能将吸收的光能传递给叶绿素a, 是光合作用不可少的光合色素,是对叶绿素捕获光能的补充。为吸收和传递光能,保护叶绿素,类胡萝卜素在植物的光合作用中具有帮助敛光,防止破坏性光氧化的作用;没有类胡萝卜素,植物几乎不能在有氧环境中进行光合作用。本研究结果表明:虫伤降低了叶绿素a、叶绿素b的含量从而影响了植物的光合作用,低密度为害对类胡萝卜素没有影响,但高密度为害导致类胡萝卜素含量增加,说明植物本身对害虫的为害有一定的抗逆性反应。类似报道如张慧杰等(2006) 的研究表明:随着美洲斑潜蝇(Liriomyza sativae)幼虫潜食叶面积的增加,蓖麻(Ricinus communis)和菜豆(Phaseolus vulgaris)的光合速率、气孔导度、蒸腾速率和气孔限制值均呈总体下降的趋势,叶片中叶绿素a、b和叶绿素总量减少。
硝态氮是植物最重要的氮源。但本研究表明芒果壮铗普瘿蚊不同程度的为害对植物叶片内硝态氮含量无显著影响。
3.2 不同为害程度对植物保护酶系的影响自前人提出自由基伤害学说以来,对活性氧自由基的产生机制、作用,及生物体内对活性氧清除系统的研究越来越受到重视。活性氧自由基的危害集中体现于它对膜脂的过氧化作用,造成膜脱脂而引起膜渗漏(李建喜等,2006)。许多研究表明:POD、SOD和CAT可以清除细胞内有害的自由基(李合生,2000)。SOD和CAT能清除植物体内产生的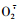
SOD可以分解
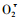
CAT存在于组织内的过氧化体中,具有催化H2O2分解为H2O与O2的作用,可使H2O2不致于与O2在铁螯合物作用下反应生成非常有害的·OH。本研究结果显示虫伤可提高植株CAT活力,这与CAT活性在抗虫品种稻株体内明显受抑制, 在感虫品种植株体内略有提高(刘裕强等,2005) 的报道相类似;也与杨振德等(2007) 报道黛袋蛾取食可诱导尾叶桉叶片内CAT活性明显升高相一致。
综上可以看出:POD,SOD和CAT活力与虫情指数呈现明显正相关,因此可以认为芒果壮铗普瘿蚊的入侵为害在不同程度上提高了植物的抗逆性。
| [] | 李赤, 于莉, 刘付东标, 等. 2007. 富贵竹中可溶性糖、蛋白质含量与细菌性茎腐病的关系. 吉林农业大学学报, 29(6): 620–622. |
| [] | 李合生. 2000. 植物生理生化实验原理和技术. 北京, 高等教育出版社: 123-185. |
| [] | 李建喜, 杨志强, 王学智. 2006. 活性氧自由基在动物机体内的生物学作用. 动物医学进展, 27(10): 33–36. DOI:10.3969/j.issn.1007-5038.2006.10.008 |
| [] | 李军, 卜文俊, 张清源. 2003. 危害芒果叶片的瘿蚊科—中国新纪录属和一新种. 动物分类学报, 28(1): 148–151. |
| [] | 李水清, 张钟宁. 2007. 松墨天牛取食和人为损伤对马尾松针叶部分化学物质含量的影响. 昆虫学报, 50(2): 95–100. |
| [] | 林振基, 蔡武煌, 吴阿松, 等. 2006. 壮铗普瘿蚊在福建省的传播为害调查. 福建农业科技(5): 27. |
| [] | 刘裕强, 江玲, 孙立宏, 等. 2005. 褐飞虱刺吸诱导的水稻一些防御性酶活性的变化. 植物生理与分子生物学报, 31(6): 643–650. |
| [] | 罗海平, 蔺万煌, 刘建丰. 2008. 水稻叶鞘3种氧化还原酶活性在褐飞虱胁迫中的变化. 植物保护, 34(5): 71–74. |
| [] | 齐慧霞, 吴学仁, 齐永顺, 等. 2005. 不同葡萄品种对霜霉病的抗性研究. 中国农学通讯, 21(4): 250–253. |
| [] | 王成霞, 董晓颖, 李培环. 2007. 桃叶片POD、SOD、CAT活性与树体矮化和生长的关系. 中国农学通报, 23(6): 353–357. |
| [] | 王琪, 严善春, 王艳军, 等. 2008. 剪叶及昆虫取食对兴安落叶松蛋白酶抑制剂的影响. 昆虫学报, 51(8): 798–803. |
| [] | 王伟新, 王宏毅. 2005. 芒果壮铗普瘿蚊生物学特性初报. 福建农业学报, 20(2): 74–76. |
| [] | 习金根, 何衍彪, 李土荣. 2006. 芒果园生草栽培主要害虫天敌分布调查. 广西热带农业, 106(5): 4–5. |
| [] | 严善春, 杨慧, 高璐璐, 等. 2009. 兴安落叶松鞘蛾对寄主挥发物的反应. 林业科学, 45(5): 94–101. DOI:10.11707/j.1001-7488.20090514 |
| [] | 杨振德, 常明山, 韦立秀, 等. 2007. 黛袋蛾取食及喷施农药对尾叶桉幼苗某些生理生化指标的影响. 东北林业大学学报, 35(7): 11–12. |
| [] | 游泳, 傅建炜, 张莉, 等. 2009. 闽台两岸芒果树冠害虫发生比较. 台湾农业探索(1): 23–28. |
| [] | 张飞萍, 尤民生. 2007. 毛竹林冠与林下层节肢动物共有种及其多样性的影响. 应用与环境生物学报, 13(4): 501–506. |
| [] | 张慧杰, 段国琪, 张战备, 等. 2006. 美洲斑潜蝇幼虫潜叶为害对几种作物光合作用的影响. 昆虫学报, 49(1): 100–105. |
| [] | Baumann D T, Bastiaans L, Goudriaan J, et al. 2002. Analysing crop yield and plant quality in an intercropping system using an eco-physiological model for interplant competition. Agriculture Systems, 73(2): 173–203. DOI:10.1016/S0308-521X(01)00084-1 |
| [] | Nykänen H, Koricheva J. 2004. Damage induced changes in woody plants and their effects on insect herbivore performance: a meta analysis. Oikos, 104(2): 247–268. DOI:10.1111/oik.2004.104.issue-2 |
| [] | Ohgushi T. 2005. Indirect interaction webs: herbivore induced effects through trait change in plants. Annual Review of Ecology, Evolution and Systematics, 36: 81–105. DOI:10.1146/annurev.ecolsys.36.091704.175523 |