 2012, Vol. 48
2012, Vol. 48文章信息
- 李荣, 张文辉, 何景峰, 周建云
- Li Rong, Zhang Wenhui, He Jingfeng, Zhou Jianyun
- 辽东栎伐桩萌苗的发育规律
- Regeneration and Development of Stump Sprouts of Quercus wutaishanica
- 林业科学, 2012, 48(3): 82-87.
- Scientia Silvae Sinicae, 2012, 48(3): 82-87.
-
文章历史
- 收稿日期:2010-10-11
- 修回日期:2011-02-18
-
作者相关文章



2. 石河子大学农学院 石河子 832000
2. 2 Agriculture College of Shihezi University Shihezi 832000
阔叶树种萌芽更新,具有迅速覆盖林地、防止土壤流失、缩短更新时间等特点,已经成为伐后更新的重要方式之一(伊力塔等,2007)。目前,国内外对阔叶林伐桩萌芽更新的研究主要集中在萌生枝的生物生态学特性(于硕等,2009)、萌生枝发生的机制及萌芽更新生长状况(李景文等,2000;2005;闫恩荣等,2005;李小双等,2007;陈少雄等,2009;Krumbiegel, 1999;王希华等,2004;Konstantinidis et al., 2006;赵睿等,2009)等方面。
辽东栎(Quercus liaotungensis)林是黄土高原地区典型地带性植被,对当地的生态防护和天然林持续发育具有举足轻重的作用(康永祥等,2007)。辽东栎伐桩有很强的萌芽能力,国内对其伐桩的研究,主要是萌芽更新、萌蘖调控等方面(伊力塔等,2007;孟令彬等,2006;2007),对于辽东栎伐桩萌苗自然存活与生长状况的研究不多,尤其在黄土高原地区,间伐后如何利用辽东栎伐桩进行更新的相关研究还未见报道,而这些正是辽东栎林采伐后进一步抚育管理的核心问题。
为此,本文以陕西黄龙山林区天然辽东栎林内的辽东栎伐桩为研究对象,通过分析间伐强度分别为13.4%和30.0%后辽东栎伐桩萌苗规律和萌生枝的存活与生长状况,以期为我国黄土高原辽东栎林采伐经营与伐后管理提供依据。
1 研究区概况与研究方法 1.1 研究区概况研究区域选择在中国西北部黄土高原地区陕西黄龙山林区。地理位置为109°38′49″—110°12′47″E, 35°28′46″—36°02′01″N,海拔962.6~1 783.5 m,属于暖温带半湿润与半干旱气候的过渡地带,年平均气温8.6 ℃,最高气温36.7 ℃,最低气温-22.5 ℃,无霜期126~186天,年均降水量611.8 mm。地带性植被为暖温带落叶阔叶林,主要乔木树种有辽东栎、油松(Pinus tabulaeformis)、茶条槭(Acer ginnala)和白桦(Betula platyphylla)等,天然植被覆盖率85%(康永祥等,2007)。目前,该地区大部分辽东栎天然次生林已达到间伐年龄(50年左右),乔木层郁闭度高达0.80~0.95,林下有树木自然枯死现象。
2004年底,选择占成树总数80%以上的辽东栎天然次生林,设计间伐材积强度为13.4%和30.0%,即弱度间伐和强度间伐(沈国舫,2009)各10.0 hm2进行间伐作业,其中126林班共标记林木6 226株(高度≥5.0 m),蓄积量860 m3,采伐材积强度13.4%,采伐蓄积量115 m3;142林班共标记林木5 893株,蓄积量490 m3,间伐材积强度30.0%,采伐蓄积量147 m3。
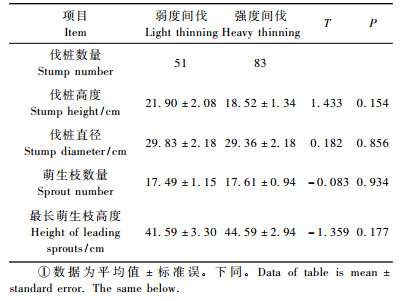
1.2 研究方法 1.2.1 伐桩萌苗调查2005年8月(间伐第1年),分别在弱度和强度间伐区,根据采伐作业路线从山底到山顶设计长250 m、宽20.0 m的2块平行样地,样地内所有辽东栎伐桩(基径≥10 cm)挂牌做永久标记。实际调查辽东栎伐桩共134株,其中弱度间伐区51株、强度间伐区83株,调查辽东栎伐桩基径、高度、萌生枝数量、最长萌生枝的高度。
2005—2010年,每年8月复查1次,调查存活伐桩(有活萌生枝)数量、每个伐桩萌生枝数量、最长萌生枝高度,统计出每年伐桩萌生枝的存活率和每年最长萌生枝的高生长量。
1.2.2 数据分析弱度间伐和强度间伐之间数据进行独立样本T检验(independent-samples T test), 不同变量间的关系进行回归模拟(regression),先采用线性回归、二次抛物线、增长曲线、对数曲线、三次抛物线、逆函数等进行回归模拟,根据显著性检验的F值和概率P值、决定系数R2等统计量,以R2值最高作为主要依据,结合每个参数有合理的生物解释,选择最优模型进行预测分析。显著性水平α设定为0.05。
统计分析全部采用SPSS11.5软件进行。
2 结果与分析 2.1 辽东栎伐桩萌苗规律 2.1.1 不同采伐强度下辽东栎的伐桩特性和伐桩萌芽力间伐第1年(2005年8月),辽东栎伐桩均有萌生枝发生,研究不同采伐强度对辽东栎的伐桩特性(伐桩高度和直径)和伐桩萌芽力(萌生枝数量和最长萌生枝高度)影响。由表 1可以看出:伐桩高度与伐桩基径在弱度采伐区略高,但与强度采伐区无显著差异;伐桩萌生枝数量与最长萌生枝高度在强度采伐区略高,但与弱度采伐区无显著差异。由此可见,伐后第1年,辽东栎伐桩萌生枝数量与最长萌生枝高度不受采伐强度影响。
|
|
研究伐桩特性(伐桩高度、直径)对伐桩萌生枝数量的影响。辽东栎伐桩高度在5.0~65.0 cm之间,伐桩萌生枝数量3~41。伐桩高度对萌生枝数量的影响表现比较复杂,当伐桩高度较低时,随着伐桩高度的增加,萌生枝数量也增加;高度达到20 cm后,萌生枝数量随着伐桩高度的增加而略有减少;高度达到50 cm后,萌生枝数量再次增加,呈三次抛物线关系(图 1)。

|
图 1 2005年伐桩高度对伐桩萌生枝数量的影响 Fig.1 Effect of the stump height on sprout number in 2005 |
辽东栎伐桩直径10~68 cm,对其与伐桩萌生枝数量进行分析,二者存在二次抛物线关系(图 2)。由此可见,随着直径的增加,辽东栎萌生枝数量先增多后减少,基径30~50 cm的萌生枝数量最多。

|
图 2 2005年伐桩直径对伐桩萌生枝数量的影响 Fig.2 Effect of the stump diameter on sprout number in 2005 |
辽东栎伐桩最长萌生枝高度31.0~141.8 cm,对伐桩萌生枝数量与最长萌生枝高度进行回归模拟,呈显著线性负相关(图 3)。由此可见,在一定范围内,随着伐桩萌生枝数量的增加,辽东栎最长萌生枝高度下降。

|
图 3 2005年伐桩萌生枝数量对最长萌生枝高度的影响 Fig.3 Effect of sprout number on height of leading sprout in 2005 |
具有存活萌生枝的伐桩占伐桩总数的比率即为伐桩存活率。调查发现:2005年辽东栎伐桩134株,均有活的萌生枝,到2010年,只有4个伐桩死亡(伐桩上萌生枝全部死亡),辽东栎伐桩存活率很高(97.01%),在弱度和强度间伐区分别为96.08%和98.80%,且不受间伐强度影响。实际调查发现:死亡的4个伐桩基径均在10~15 cm之间,未采伐前树势衰弱,伐后第1年每桩萌发3~4个萌生枝,新生枝条高20~30 cm,生长细弱,到第3~4年全部萌生枝干枯而导致伐桩死亡。
2.2.2 辽东栎伐桩最长萌生枝高度变化伐后6年期间,辽东栎伐桩最长萌生枝高度在不同间伐强度间的生长趋势相同,始终呈现出弱度<强度的趋势(图 4),伐后第1年,伐桩最长萌生枝高度平均为(43.45±1.13)cm,不同强度间无显著差异(P=0.197),随后2~4年分别为(52.52±1.28)cm(P=0.093)、(67.28±1.57)cm(P=0.072)和(74.40±1.62)cm(P=0.067),到间伐5~6年,不同强度间存在显著差异,分别为(83.41±1.41)cm(P=0.031)和(89.17±1.46)cm,(P=0.032),说明间伐强度对后期辽东栎伐桩最长萌生枝高度有显著影响。

|
图 4 伐后6年期间伐桩最长萌生枝高度变化 Fig.4 Change of the height of leading sprout through time over 6 years |
间伐后1~6年,辽东栎伐桩最长萌生枝平均高度逐年递增(图 4),但是平均每年的高生长量不同。第1年萌生枝生长最快(43.45 cm),第2年下降到9.07 cm,仅仅是第1年的20.87%,第3年较高(14.76 cm),随后4~6年依次为7.12,9.01和5.76 cm。由此可见,间伐后,辽东栎伐桩萌生枝迅速生长,而后逐步下降。
2.3 辽东栎伐桩萌生枝的数量和数量结构变化 2.3.1 辽东栎伐桩萌生枝的数量变化伐后6年期间,辽东栎伐桩萌生枝数量在不同间伐强度间无显著差异,伐后第1年伐桩萌生枝数量平均为17.57±0.73(P=0.934),到第2年下降到15.40±0.70(P=0.717),随后3~6年分别为10.88±0.52(P=0.956)、8.72±0.45(P=0.258),7.49±0.38(P=0.931)和6.95±0.32(P=0.158)(图 5)。由此可见,调查期间,伐桩萌生枝存在自我稀疏过程且萌生枝数量始终不受间伐强度影响。伐后第5~6年,伐桩萌生枝数量下降幅度缓慢,说明该期伐桩萌生枝数量趋于稳定。

|
图 5 伐后6年期间伐桩萌生枝数量变化 Fig.5 Change of the sprout number through time over 6 years |
伐后6年期间,辽东栎伐桩萌生枝的数量结构发生了很大变化(图 6)。2005年(伐后第1年),伐桩萌生枝的数量在3~41之间,其中10~21的伐桩数量占伐桩总数的55.22%;到2010年(伐后第6年),伐桩存活萌生枝数量仍然较高,在1~16之间,其中4~9的伐桩数量占伐桩总数的59.23%。这说明,调查期间伐桩萌生枝数量大幅度减少,且伐桩产生的萌生枝越多,后期该伐桩萌生枝死亡的数量也越多。

|
图 6 2005和2010年伐桩的萌生枝的数量结构变化 Fig.6 Number structure of sprout in 2005 and 2010 |
在黄土高原地区,辽东栎伐桩的萌芽潜力很大,伐桩高度对萌生枝数量的影响表现比较复杂,呈三次抛物线关系,这与陈少雄等(2009)研究尾叶桉U6无性系(Eucalyptus urophylla clone U6)萌芽规律一致,但与其他人研究阔叶树种的结果不一致,如荆涛等(2002)研究水曲柳(Fraxinus mandshurica)伐桩认为二者呈线性负相关;Edward等(2008)研究热带雨林4个树种伐桩认为萌枝数量与伐桩高度显著线性相关(3个正相关,1个负相关);伊力塔等(2006)研究辽东栎伐桩认为萌生枝数量随着伐桩高度的增加先增多再减少,二者呈二次抛物线关系。这种伐桩高度影响萌生枝数量表现的复杂性,分析其原因,焦点在伐桩产生萌生枝的部位上。Edward等(2008)认为萌生枝主要产生在接近伐口或者在伐口点上,因为砍伐后,伐口附近的芽比伐桩基部的芽受到更多的刺激;但在本研究中发现,辽东栎伐桩萌生枝产生的部位主要在伐桩基部,其次是在接近伐口的部位。萌生枝产生部位的不同,是否与物种的特性或物种所处的发育阶段有关,需要做进一步的研究。
辽东栎伐桩萌生枝数量随着伐桩直径的增大呈先增多后减少的规律,这与伊力塔等(2006)研究辽东栎的结论一致,在其他栎类研究中也有相类似的报道(Tworkoski et al., 1990;Groninger et al., 1998;Weigel et al., 2002)。伊力塔等(2006)研究认为树木采伐后强烈刺激了伐桩上的休眠芽和不定芽萌发产生萌生枝,因此,萌生枝数量与休眠芽和不定芽的数量有关。随着树木的生长,由原生分生组织形成休眠芽的数量不变,由次生分生组织所产生的不定芽却随着树木直径的增大而增多;但增长到一定程度后,由于树木自身开花结实及老化导致其径分生能力下降,产生不定芽的能力下降(李景文等,2000;2005)。基径在30~50 cm(处于青年期)的伐桩萌生枝数量最多,说明此期间辽东栎个体碳水化合物和其他营养物质的储备最多、分生能力最强。
辽东栎伐桩萌生枝数量影响最长萌生枝高度,二者呈显著线性负相关,这种观点在辽东栎上没有相关报道,但在McLaren等(2003)研究热带干旱区森林7个阔叶树种、Edward等(2008)研究热带雨林5个阔叶树种中有类似报道。这种研究结果可以用Midgley(1996)提出的“萌生枝数量——高度权衡”假说来解释。Midgley提出萌芽生长导致高度生长代价这一观点,认为产生较多萌生枝虽然可提高更新的成功率,但是萌生枝生长需要消耗大量的资源,如果过多的资源用于萌生枝生长,即伐桩萌生枝数量增加,可能导致高度生长的投入减少,即萌生枝高度生长降低。
3.2 辽东栎伐桩萌生枝的生长调查期间,伐桩萌生枝高度逐步递增,但每年的生长量不同。间伐后辽东栎伐桩萌生枝迅速生长,而后萌生枝每年高生长量逐步下降,这种趋势与其他栎类研究相类似(Cobb et al., 1985;Gardiner et al., 1997;Lockhart et al., 2007),主要是随着时间推移,伐桩内的养分(主要是储存的碳水化合物和激素物质)大量减少而影响萌生枝生长。Cobb等(1985)还认为,早期伐桩萌生枝迅速生长与随后略微下降与伐桩萌生枝之间和林地内上层植物与下层植物之间的竞争有关。
3.3 辽东栎伐桩存在“萌代主”现象萌代主分析萌芽更新的意义关键在于萌生枝能否代替被采伐的主干形成大树,即“萌代主”的问题(李景文等,2005)。伐后1~6年,辽东栎伐桩的每桩萌生枝数量在逐步下降,这与Lockhart等(2007)研究宝塔栎(Quercus pagoda)趋势一致,他认为这是栎类在生长过程中对养分的强烈竞争而造成的自我稀疏过程。研究辽东栎伐桩2005和2010年萌生枝的数量结构变化,发现辽东栎伐桩产生萌生枝的能力很强,导致随后萌生枝之间的竞争激烈、伐桩萌生枝数量大幅度减少,说明辽东栎伐桩的萌生枝存在“萌代主”现象;但伐后第6年存活伐桩萌生枝数量仍然较高,且实际调查发现,保留下来的萌生枝之间的长度和基径差异不大。伐桩萌生枝长势均衡且灌丛状生长,从长远来看,不利于辽东栎林的更新。
自然状态下,辽东栎伐桩萌生枝之间存在强烈的自疏过程,但“萌代主”萌生枝保留的数量过多,萌生的大树干形不良,即所谓“砍刀形”大树,木材利用价值降低(李景文等,2005)。因此,辽东栎在伐桩萌芽更新过程中,前期应进行人为除蘖定株,否则,萌生枝过多,存在“萌生枝数量——高度权衡”现象,且后期竞争激烈,影响萌生枝的生长。孟令彬等(2006;2007)研究认为每个辽东栎伐桩应该保留2~3个生长良好的萌生枝为宜。
3.4 不同强度间伐对辽东栎伐桩的萌芽力与生长的影响辽东栎采伐后当年均有萌生枝发生,且采伐当年伐桩萌生枝数量与最长萌生枝高度不受采伐强度影响,这与其他栎类相类似(Gardiner et al., 1997;Lockhart et al., 2007)。伐后第6年,强度采伐区辽东栎伐桩最长萌生枝高度年生长量远远高于弱度采伐区,这与Gardiner等(1997)研究伐后5年的黑栎(Quercus nigra)一致;但是,Lockhart等(2007)研究伐后4年的宝塔栎认为,采伐强度不会影响萌生枝高度。这2种不同的结论,可能由于研究的物种生物学特性以及立地条件不同所导致的。
本研究认为影响辽东栎伐桩萌生枝高度生长的环境因素主要是光照。萌生枝的高度生长在阳光较充足的微生境下(强度间伐)比荫蔽的树冠微生境下(弱度间伐)高,因为前期高度的迅速生长有利于萌生枝的存活和生长,这与贺顺钦等(2001)认为光照强度对辽东栎幼苗生长和存活有较大影响的研究结论类似。
4 结论在黄土高原地区,辽东栎伐桩的萌芽潜力很大,伐桩高度影响伐桩萌生枝数量,二者呈三次抛物线关系。考虑到资源的节约利用,间伐辽东栎时,伐桩的高度应尽量低于20 cm。
自然状态下,辽东栎伐桩直径影响伐桩萌生枝数量,且萌生枝数量随着伐桩直径的增大呈先增多后减少的规律。直径30~50 cm的辽东栎萌生枝数量最多,是进行伐桩无性更新中的重点经营对象。
辽东栎伐桩前期存在“萌生枝数量——高度权衡”现象,在生长过程中存在强烈的自疏过程,后期存在“萌代主”现象,因此,间伐前期伐桩要及时摘芽除蘖,这样有利于萌生枝的高度生长。
影响辽东栎伐桩萌生苗高度生长的主要因素是光照,因此,强度间伐(间伐强度30.0%)更有利于辽东栎伐桩的生长与更新。
| [] | 陈少雄, 李天会, 李志辉, 等. 2009. 尾叶桉U6无性系萌芽性能研究. 林业科学研究, 22(5): 657–661. |
| [] | 贺顺钦, 王发其. 2001. 辽东栎苗木早期生长与光的关系. 林业科学研究, 14(6): 697–700. |
| [] | 荆涛, 马万里, JoniKujansuu, 等. 2002. 水曲柳萌芽更新的研究. 北京林业大学学报, 24(4): 12–15. |
| [] | 康永祥, 岳军伟, 张巧明. 2007. 黄龙山林区辽东栎群落类型划分及其生物多样性研究. 西北林学院学报, 22(3): 7–10. |
| [] | 李景文, 刘世英, 王清海, 等. 2000. 三江平原低山丘陵区水曲柳无性更新研究. 植物研究, 20(2): 215–220. |
| [] | 李景文, 聂绍荃, 安滨河. 2005. 东北东部林区次生林主要阔叶树种的萌芽更新规律. 林业科学, 41(6): 72–77. DOI:10.11707/j.1001-7488.20050611 |
| [] | 李小双, 彭明春, 党承林. 2007. 植物自然更新研究进展. 生态学杂志, 26(12): 2081–2088. |
| [] | 孟令彬, 包维楷, 庞学勇, 等. 2006. 萌蘖调控对辽东栎留存萌生株生长与结实的影响. 应用生态学报, 17(10): 1771–1776. DOI:10.3321/j.issn:1001-9332.2006.10.001 |
| [] | 孟令彬, 包维楷, 庞学勇, 等. 2007. 萌蘖调控对辽东栎再萌生能力的影响. 应用与环境生物学报, 13(1): 18–22. |
| [] | 沈国舫. 2009. 森林培育学. 北京, 中国林业出版社: 303-304. |
| [] | 王希华, 严晓, 闫恩荣, 等. 2004. 天童几种常绿阔叶林优势种在砍伐后萌枝更新的初步研究. 武汉植物学研究, 22(1): 52–57. |
| [] | 闫恩荣, 王希华, 施家月, 等. 2005. 木本植物萌枝生态学研究进展. 应用生态学报, 16(12): 2459–2464. DOI:10.3321/j.issn:1001-9332.2005.12.046 |
| [] | 伊力塔, 韩海荣, 马钦彦, 等. 2006. 灵空山辽东栎萌芽更新的灰色关联分析. 山西林业科技(1): 23–25. |
| [] | 伊力塔, 韩海荣. 2007. 山西灵空山林区辽东栎萌芽更新规律研究. 林业资源管理(4): 57–61. |
| [] | 于硕, 王蝾, 刘敏, 等. 2009. 天目山米心水青冈群落物种组成特点与萌条更新. 生态学杂志, 28(2): 182–187. |
| [] | 赵睿, 周学峰, 徐娜娜, 等. 2009. 米心水青冈种群萌条更新与高度生长. 生态学报, 29(7): 2665–2669. |
| [] | Cobb S W, Miller A E, Zahner R. 1985. Recurrent shoot flushes in scarlet oak stump sprouts. For Sci, 31(3): 725–730. |
| [] | Edward N M, Ed T F. 2008. Sprouting of woody species following cutting and tree-fall in a lowland semi-deciduous tropical rainforest, North-Western Uganda. Forest Ecology and Management, 255(3/4): 982–992. |
| [] | Gardiner E S, Helmig L M. 1997. Development of water oak stump sprouts under a partial overstory. New Forest, 14(1): 55–62. DOI:10.1023/A:1006502107495 |
| [] | Groninger J W, Stein H D, Zedaker S M, et al. 1998. Growth response and cost comparisons for precommercial thinning methods of Appalachian oak stump sprout clumps. South J Appl For, 22(1): 19–23. |
| [] | Konstantinidis P, Tsiourlis G, Xofis P. 2006. Effect of fire season, aspect and pre-fire plant size on the growth of Arbutus unedo L. (strawberry tree) resprouts. Forest Ecology and Management, 225(1/3): 359–367. |
| [] | Krumbiegel A. 1999. Growth forms of biennial and plurennial vascular plants in central Europe. Nordic Journal of Botany, 19(2): 217–226. DOI:10.1111/j.1756-1051.1999.tb00667.x |
| [] | Lockhart B R, Chambers J L. 2007. Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley, USA. New Forests, 33(2): 183–192. DOI:10.1007/s11056-006-9022-7 |
| [] | McLaren K P, McDonald M A. 2003. Coppice regrowth in a disturbed tropical dry limestone forest in Jamaica. Forest Ecology and Management, 180(1/3): 99–111. |
| [] | Midgley J J. 1996. Why the world vegetation is not totally dominated by resprouting plants: because resprouters are shorter than reseeders. Ecography, 19(1): 92–94. DOI:10.1111/eco.1996.19.issue-1 |
| [] | Steven D D, Wright S J. 2002. Consequences of variable reproduction for seedling recruitment in three neotropical tree species. Ecology, 83(8): 2315–2327. DOI:10.1890/0012-9658(2002)083[2315:COVRFS]2.0.CO;2 |
| [] | Tworkoski T J, Ross M S, Hopper G M. 1990. Analysis of chestnut and scarlet oak stump sprouts growth. Can J For Res, 20(1): 112–116. DOI:10.1139/x90-016 |
| [] | Weigel D R, Peng C J. 2002. Predicting stump sprouting and competitive success of five oak species in southern Indiana. Can J For Res, 32(4): 703–712. DOI:10.1139/x02-042 |