 2012, Vol. 48
2012, Vol. 48文章信息
- 宋金凤, 李锋, 王亚军, 高红松, 崔晓阳, 彭红梅
- Song Jinfeng, Li Feng, Wang Yajun, Gao Hongsong, Cui Xiaoyang, Peng Hongmei
- 低温胁迫下外源有机酸对长白落叶松幼苗生理生化特性的影响
- Effects of Exogenous Organic acids on Physiological and Biochemical Characteristics of Larix olgensis seedlings under Low Temperature Stress
- 林业科学, 2012, 48(2): 89-95.
- Scientia Silvae Sinicae, 2012, 48(2): 89-95.
-
文章历史
- 收稿日期:2011-03-15
- 修回日期:2011-09-27
-
作者相关文章



在温带林区,植物常遭受春季夜间的冷害侵扰,此时植物膜脂物相改变,膜结构遭到破坏、透性增加,细胞超微结构明显变化,细胞内发生一系列生理和代谢功能紊乱,如叶绿素合成受抑、叶绿体结构被破坏、光合速率降低、有机物运输受阻、活性氧大量产生、引起细胞氧化胁迫(Prasad et al., 1994),最终导致生产力降低。长白落叶松(Larix olgensis)是我国东北高寒地区生长快、适应性强的主要用材树种之一,又因其冠形美观、抗烟力强,还是一个优良的园林绿化树种,目前其造林面积逐年扩大。在我国东北林区,长白落叶松育苗常在春季进行,一般将种子催芽后在4月末至5月初播种(李冬梅等,2008),由于东北林区气候寒冷,长白落叶松幼苗常遭受冷害侵扰,特别是在夜间伤害更重,导致其生长缓慢,因此找到提高长白落叶松幼苗抗寒性的有效方法、延长苗木的生长期是当前生产急需解决的问题之一。
有机酸是森林生态系统中普遍存在的一类高分子有机物,对植物抗寒性、抗旱性、抗重金属胁迫、抗养分胁迫等多种抗性的形成具有重要作用(Oburger et al., 2009; Schmitt et al., 2011)。低温胁迫下,外源水杨酸(SA)等有机酸能通过多种机制有效提高植物细胞膜结构的稳定性,增强叶片抗氧化胁迫的能力,最终缓解低温胁迫对植物的伤害,这已在水曲柳(Fraxinus mandshurica)、黄瓜(Cucumis sativus)、西瓜(Citrullus lanatus)、山桃(Amygdalus davidiana)、水稻(Oryza saliva)等多种植物的抗寒性试验中得以证实,且供试植物品种、有机酸种类和处理方式不同,抗寒性提高的程度和有机酸适宜浓度也不同(Tsuchidaa et al., 2010; 吴楚等,2002a)。另外,低温等逆境因子还诱导植物根系特异性地分泌某些有机酸,这是一种较为普遍的主动适应性反应(Tolrá et al., 2005; Zeng et al., 2008)。可以假设,外源有机酸亦能通过多种机制提高长白落叶松幼苗对低温伤害的抵御能力,进而提高其生产力水平。本文选择草酸、柠檬酸和苹果酸3种我国东北林区代表性森林凋落物中的优势有机酸(宋金凤等,2004),通过模拟自然低温,在长白落叶松幼苗转入4℃低温前,用不同浓度上述有机酸溶液进行叶面喷施处理,研究外源有机酸对长白落叶松幼苗受低温胁迫后多种生理生化特性的影响,包括细胞膜透性、多种抗氧化酶活性、膜脂过氧化产物丙二醛含量和渗透调节物质含量等,以探讨利用有机酸提高长白落叶松幼苗抗寒能力的可行性及机制,研究对于了解天然更新过程中长白落叶松幼苗抗寒能力的形成机制具有重要的理论意义,对育苗实践中长白落叶松幼苗抗寒能力的提高也具有指导作用。
1 材料与方法 1.1 试验材料和处理方法试验在东北林业大学帽儿山实验林场温室内进行。长白落叶松种子经过选种、消毒后,以雪藏法进行催芽,4月初播种,覆土料为质地均一、无杂质的A1层暗棕壤。育苗在营养钵(底径16.2cm、上口径18.4cm、高20.0cm)内进行,每钵播种20粒,钵上沿土壤空出2~3cm,以便浇水和有机酸处理。5月下旬,拔除营养钵内部分幼苗,每钵仅留10株,再在土壤表层洒一层沙子以减少水分蒸发。苗木在温室内正常光照和浇水,缓苗半月后,于6月初进行低温和有机酸处理,此时幼苗平均苗高6.5cm,平均基径1.34cm。
用喷壶向长白落叶松幼苗的叶片上、下表面喷施有机酸溶液(分别为草酸、柠檬酸和苹果酸溶液,浓度分别为0,0.2,1.0,5.0,25.0和50.0mmol·L-1,均以有机钠盐形式喷施),至叶面均匀湿润、向下滴液体为止。每天晚上6:00喷施,连喷3天,第3天有机酸喷施2h后,将苗木连同营养钵一起放入4℃冰箱中低温处理12h,取出苗木在室温下恢复24h后取样分析。对照为室温(23℃)、喷施蒸馏水处理(以CK表示)。每处理10钵。
1.2 样品采集与测定随机采集足够量的第11~20片成熟真叶(幼苗中部),立即测定相对电导率、丙二醛(MDA)、脯氨酸(Pro)、可溶性蛋白和叶绿素含量,以及超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)和抗坏血酸过氧化物酶(APX)活性,其中相对电导率用上海雷磁DDS-6700电导仪测定; MDA含量采用硫代巴比妥酸(TBA)比色法; Pro含量采用酸性茚三酮法; 可溶性蛋白含量采用考马斯亮蓝G-250染色法; 叶绿素含量采用丙酮提取分光光度法; SOD活性采用氮蓝四唑(NBT)光化还原法; POD活性采用愈创木酚法; CAT活性采用紫外吸收法; APX活性采用紫外吸收法(李合生,2000)。每处理重复3次。
2 结果与分析 2.1 有机酸对低温下长白落叶松叶片细胞膜系统的保护作用 2.1.1 膜渗透性变化4℃低温增大了长白落叶松幼苗的细胞膜透性,叶片相对电导率略高于常温对照。外源有机酸处理后,膜透性变化显著:在0.2~5.0mmol·L-1较低浓度内,相对电导率均下降,且随有机酸浓度增加而降低; 有机酸浓度再高,不同有机酸又表现出不同的变化趋势,大多数处理均高于低温无有机酸处理,说明有机酸浓度过高对抑制苗木电解质渗出起反作用。柠檬酸对降低细胞膜透性的作用最强,苹果酸其次,草酸较弱(图 1)。

|
图 1 有机酸对长白落叶松幼苗叶片相对电导率的影响
Fig.1 Effects of organic acids on relative conductivity in L. olgensis seedlings
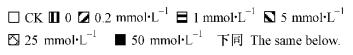 不同字母表示差异显著(P<0. 05)。下同。
Bars labeled with different letters were significantly different (P < 0. 05).The same below. 不同字母表示差异显著(P<0. 05)。下同。
Bars labeled with different letters were significantly different (P < 0. 05).The same below.
|
经4℃处理后,长白落叶松幼苗MDA含量增加,增幅为40.51%,说明低温胁迫已使落叶松幼苗的细胞膜受到损伤,抗氧化能力降低。有机酸处理后MDA含量降低,说明膜系统的过氧化损伤得以恢复,从而有效抵御了低温对苗木的伤害。草酸、柠檬酸和苹果酸分别以5.0,1.0和1.0mmol·L-1处理效果最佳,降低幅度顺序为:柠檬酸>苹果酸>草酸(图 2)。

|
图 2 有机酸对长白落叶松幼苗叶片MDA含量的影响 Fig.2 Effects of organic acids on MDA contents in L. olgensis seedlings |
与对照相比,低温处理后,未经有机酸处理的苗木SOD,POD和APX活性均略有升高,说明苗木能通过自身的酶促保护机制抵御低温伤害。但4℃低温显著降低CAT活性,说明低温胁迫后氧自由基的产生速率可能超过了系统的清除能力,细胞受到伤害。0.2~50.0mmol·L-1有机酸增强了低温下苗木的SOD活性,且SOD对有机酸反应敏感,0.2mmol·L-1低浓度时即产生明显效果。SOD活性与有机酸浓度间大体呈峰型曲线变化,即先升后降,但一直高于未经有机酸处理的幼苗。所有浓度有机酸处理还增加了POD和CAT活性,大多数浓度也增加了APX活性,其中POD和APX活性均以1.0mmol·L-1增幅最大,CAT活性则在1.0或5.0mmol·L-1达峰值,增幅均达到显著水平。3种有机酸诱导上述4种抗氧化酶活性增加的程度为:柠檬酸>苹果酸>草酸,大多数浓度处理时都较明显(图 3~6)。

|
图 3 有机酸对长白落叶松幼苗叶片SOD活性的影响 Fig.3 Effects of organic acids on SOD activity in Larix olgensis seedlings |

|
图 4 有机酸对长白落叶松幼苗叶片POD活性的影响 Fig.4 Effects of organic acids on POD activity in L. olgensis seedlings |

|
图 5 有机酸对长白落叶松幼苗叶片APX活性的影响 Fig.5 Effects of organic acids on APX activity in L. olgensis seedlings |

|
图 6 有机酸对长白落叶松幼苗叶片CAT活性的影响 Fig.6 Effects of organic acids on CAT activity in L. olgensis seedlings |
Pro是植物体内最有效的渗透调节物质之一,在低温胁迫下能增加植物组织的束缚水含量、降低细胞水势、增强持水力,从而提高植物的抗寒性。可溶性蛋白也是一种重要的渗透调节物质,低温能引起植物体内的代谢改变,进而蛋白质发生变化,主要表现在可溶性蛋白含量变化及产生抗冷蛋白。低温处理后,落叶松幼苗Pro含量比常温对照增加了5.17%,说明低温胁迫后苗木能自发产生某种适应机制,诱导Pro积累以降低低温危害,但可溶性蛋白含量却下降5.30%。3种有机酸处理后,Pro含量均高于未加有机酸组,其中以5.0mmol·L-1浓度增效最明显,这说明,适宜浓度的有机酸在低温诱导的Pro积累中发挥积极作用,对提高苗木的低温胁迫适应性效果明显(图 7)。与Pro含量的变化类似,在0.2~5.0mmol·L-1浓度内,可溶性蛋白含量随有机酸浓度增加大致呈线性增加,5.0mmol·L-1时达峰值,浓度再大则下降,但仍高于对照。3种有机酸对Pro和可溶性蛋白含量的增加顺序均为:柠檬酸>苹果酸>草酸(图 8)。

|
图 7 有机酸对长白落叶松幼苗叶片Pro含量的影响 Fig.7 Effects of organic acids on Pro contents in L. olgensis seedlings |

|
图 8 有机酸对长白落叶松幼苗叶片可溶性蛋白含量的影响 Fig.8 Effects of organic acids on soluble protein contents in L. olgensis seedlings |
长白落叶松幼苗对低温较敏感,遭受4℃低温后叶绿素含量迅速降低,包括叶绿素a、叶绿素b、类胡萝卜素含量和叶绿素总量,叶绿素a/b也下降。不同浓度有机酸处理能减缓长白落叶松幼苗叶绿素的下降趋势,与无有机酸处理相比,大多数处理差异显著,其中以5.0mmol·L-1浓度效果最好。从有机酸处理后叶绿素a/b变化看,大多数处理表现为下降趋势。3种有机酸对叶绿素含量的影响强弱为:柠檬酸>苹果酸>草酸,即同浓度3种有机酸处理时,叶绿素的降低程度为:草酸>苹果酸>柠檬酸,这在大多数浓度处理时均有体现(表 1)。
|
|

细胞膜结构的稳定性和MDA含量能反映植物受低温伤害后细胞膜脂过氧化程度的强弱(梁建萍等,2007; Campos et al., 2003),电解质渗透率和MDA含量越高,植物受伤害越重(吴锦程等,2010)。低温胁迫下,有机酸显著降低长白落叶松幼苗的相对电导率和MDA含量,说明有机酸能增强苗木的抗低温能力,这与前人结果一致(吴楚等,2002b;吕军芬等,2004; 蔺经等,2004),但本文与前人研究中有机酸处理的最佳浓度不同(蔺经等,2004;吕军芬等,2004; 黄爱霞等,2003),这可能与植物和有机酸种类不同有关。
SOD是最重要的清除氧自由基的酶之一,主要功能是催化超氧阴离子自由基发生歧化反应生成H2O2,还可通过化学反应清除单线态氧和羟自由基(吴楚等,2002a; Noctor et al., 1998)。低温下植物体内SOD活性的变化因研究对象和处理方法不同而异(周艳虹等,2003; 胡文海等,2003)。本研究发现:低温诱导长白落叶松SOD活性提高,表明其为避免遭受低温伤害已做出适应性反应,也说明苗木在逆境下已生成了相应的氧自由基,氧自由基的产生可能已超过SOD等抗氧化系统的清除能力,故部分未能清除的活性氧可能已引起膜脂过氧化,导致膜系统损伤(相对电导率和MDA含量均增加)。很多研究发现:SA等有机酸诱导SOD活性增加从而清除活性氧(Scandalio,1997),但也有相反的结果(吴楚等,2002b)。有机酸处理后,长白落叶松幼苗一直保持较高的SOD活性,说明有机酸能提高苗木对低温伤害的抵御能力,这可能与有机酸增强土壤和植物体内的养分供应有关。具体讲,有机酸显著促进土壤中P,Fe等养分的活化释放,并促进植物的吸收(Pizzeghello et al., 2006; Palomo et al., 2006;宋金凤等,2008),因此,有机酸可能间接提高植物的生理活动过程,进而促进酶的合成。3种有机酸对暗棕壤P,Fe等养分活化的程度为:柠檬酸>苹果酸>草酸(宋金凤等,2008),对SOD活性的提高程度也如此,这主要与3种有机酸的化学结构、离解常数及有机酸配体和金属形成络合物的稳定性大小有关。3种有机酸对落叶松叶片内POD,CAT和APX等其他保护酶活性及可溶性蛋白、Pro等含量的影响规律与此一致。
POD在逆境下酶保护系统中的作用相对较弱,但其底物不十分严格,在保护膜结构中仍起一定作用。前人研究发现:SA等有机酸诱导水曲柳、番茄(Lycopersicon esculentum cv.‘Zhongza No.9’)、苜蓿(Medicago sativa)、山桃、水稻等植物体内POD活性增加,从而有利于植物迅速清除活性氧(吴楚等,2002b;张彩峡,2010; 张蕊,2006)。本研究结果显示:有机酸还诱导新的POD同工酶(李兆亮等,1998)。CAT能催化H2O2转变为H2O,其活性越高,植物耐寒能力越强。低温使长白落叶松幼苗CAT活性下降,这与前人研究一致(张彩峡,2010; 胡文海等,2003)。APX直接负责清除叶绿体和溶质中的H2O2(吴楚等,2002a; 张蕊,2006),一般低温胁迫后APX活性增加(贾庆虎,2007),本文结果与此相同。外源有机酸对植物体内CAT和APX活性的影响,不同研究者针对不同植物和有机酸得到的结论不同(Dat et al., 2000; 吴楚等,2002b)。关于SA提高植物抗氧化酶活性和抗胁迫能力的机制,可能由于植物体内SA受体蛋白基因与过氧化物酶基因高度同源,外源SA进入体内能激活SOD酶活性; SA还参与植物体内茉莉酸代谢,后者能提高胁迫下植物体内SOD,POD和CAT等酶的活性,而SOD和POD主要功能是消除胁迫诱导产生的活性氧自由基,抑制膜内不饱和脂肪酸过氧化作用及其产物MDA积累(李德红等,1995)。尽管本研究采用了与SA不同的3种有机酸,但它们与SA一样,都属于森林生态系统中普遍存在的一类低分子有机酸类物质,其以何种机制提高长白落叶松幼苗的抗氧化酶活性和抗寒性可能与SA类似。另外,有机酸还提高暗棕壤中P,Fe等养分的有效性,并促进植物吸收,进而可能促进植物代谢,间接促进抗氧化酶的合成与积累,提高酶系统的保护能力。
在无严重冷害的温度下,游离Pro含量增加是植物对低温胁迫的一种适应性生理反应(蔺经等,2004;张彩峡,2010),抗寒能力越强的植物游离Pro的增加也越多(王小华等,2008)。本研究发现:有机酸提高长白落叶松的Pro含量,表明长白落叶松可通过渗透调节而增加其对低温胁迫的适应,这与前人关于西瓜(吕军芬等,2004)、山桃(蔺经等,2004)、水稻(张蕊,2006)等研究的结果一致。同样,本文与以往研究的有机酸处理的最佳浓度也不同。前人进行有机酸处理的方式与本文有同有异,大多数进行叶面喷施或灌根,但无论处理方式如何,有机酸提高Pro含量进而增强抗寒性的结论一致。
关于低温后植物体内可溶性蛋白含量的变化,有人认为可溶性蛋白含量与抗低温胁迫间有平行增长关系,在植物冷驯化中还有新蛋白质合成(李美如等,1995); 还有人认为冷害加快蛋白质分解变性,其含量与抗寒性无关(艾希珍等,1999)。本研究发现:低温处理后长白落叶松体内的可溶性蛋白含量降低,说明低温已造成其合成受阻或者分解变性加快,这与前人关于黄瓜(贾庆虎,2007)、辣椒(Capsicum annum)(李盈,2009)、苜蓿(张彩峡,2010) 等研究并不一致,可能与植物种类不同有关。本研究还发现:有机酸处理后可溶性蛋白含量显著增加,此结果与以往研究一致(张蕊,2006; 贾庆虎,2007),说明增加蛋白质含量也是长白落叶松幼苗适应低温胁迫的重要机制之一,但是否有新蛋白质合成,还需进一步探讨。关于SA如何通过提高植物体内可溶性蛋白含量而增强其抗寒性,一方面,SA活化了细胞内某些蛋白质的合成基因,提高了蛋白质合成量; 另一方面,SA可能改变植物体内的物质代谢途径,降低低温条件下蛋白质的变性降解速度。草酸、柠檬酸、苹果酸提高长白落叶松体内可溶性蛋白含量的机制可能与SA类似。
植物叶片叶绿素含量和光合作用强弱对森林生产力的大小具有决定意义。本研究发现:有机酸减弱长白落叶松叶绿素含量的降低趋势,其机制可能包括以下2方面,通常可能是2种机制同时起作用。首先,有机酸显著提高植物体内多种酶的活性,包括硝酸还原酶(NR)、亚硝酸还原酶(NIR)、转化酶(INV)、谷氨酸合成酶(GOGAT)PAL,CAT,POD,PPO,SOD等(束良佐等,2002),有机酸可能通过提高参与叶绿素生物合成过程中酶的活性而促进叶绿素合成; 其次,矿质元素对叶绿素的形成有重要影响,其中Mg是组成叶绿素的元素,Fe,Mn,Cu,Zn等可能是叶绿素形成过程中某些酶的催化剂,有机酸能通过酸化、螯合、氧化还原反应、减少吸附等机制显著提高土壤中Fe,Mn,Cu,Zn,Mg的有效性(Li et al., 2006; Reichard et al., 2007),进而促进叶绿素合成。有机酸对叶绿素含量的影响与有机酸种类和浓度、植物种类等密切相关,有机酸对落叶松幼苗叶绿素含量的提高顺序为:柠檬酸>苹果酸>草酸,这与3种有机酸促进暗棕壤养分释放的顺序一致。目前有一些有机酸增加正常生长植物体内叶绿素含量的报道,有机酸种类涉及苹果酸、柠檬酸、酒石酸、乳酸、SA等,叶绿素包含了叶绿素a、叶绿素b和类胡萝卜素(罗毅等,2006); 胁迫条件下有机酸的添加研究也得到了类似结果(束良佐等,2002; 张蕊,2006)。有机酸减缓低温下叶绿素含量的下降趋势有一定的浓度限制,浓度过高反而起反作用(吕军芬等,2004)。SA等有机酸还有效阻止植物叶片光合速率的下降速度(康国章等,2003; 张蕊,2006)。本文未测定光合效率,但由有机酸处理后长白落叶松叶绿素的变化趋势可以推断,草酸、柠檬酸和苹果酸亦能提高苗木的净光合速率,从而促使其克服东北林区的高寒天气,延长生长期,提高产量。
总之,草酸、柠檬酸和苹果酸能提高低温胁迫下长白落叶松幼苗的SOD,POD,CAT和APX等多种保护酶活性,降低MDA含量,明显提高渗透调节物质的含量,减弱叶绿素含量的下降趋势,并最终提高苗木抗寒性,适宜的最佳使用浓度为5.0mmol·L-1。
| [] | 艾希珍, 于贤昌, 王绍辉, 等. 1999. 低温胁迫下黄瓜嫁接苗与自根苗某些物质含量的变化. 植物生理学通讯, 35(1): 26–28. |
| [] | 胡文海, 李晓红. 2003. 低温弱光对番茄叶片和根系中抗氧化酶系统的影响. 井冈山师范学院学报:自然科学, 24(5): 19–22. |
| [] | 黄爱霞, 佘小平. 2003. 水杨酸对黄瓜幼苗抗冷性的影响. 陕西师范大学学报:自然科学版, 31(3): 107–109. |
| [] | 贾庆虎. 2007. ABA对黄瓜幼苗抗冷性的影响. 山东农业大学硕士专业学位论文: 15–30. |
| [] | 康国章, 欧志英, 王正询, 等. 2003. 水杨酸诱导提高香蕉幼苗耐寒性的机制研究. 园艺学报, 30(2): 141–146. |
| [] | 李德红, 潘瑞炽. 1995. 水杨酸在植物体内的作用. 植物生理学通讯, 31(2): 144–149. |
| [] | 李冬梅, 黄春泰. 2008. 长白落叶松育苗技术. 黑龙江生态工程职业学院学报, 21(3): 31–32. |
| [] | 李合生. 2000. 植物生理生化试验原理和技术. 北京:高等教育出版社: 134–263. |
| [] | 李美如, 刘鸿先, 王以柔. 1995. 植物细胞中的抗寒物质及其与植物抗冷性的关系. 植物生理学通讯, 31(5): 324–328. |
| [] | 李盈. 2009. PEG处理对低温弱光下辣椒幼苗生理生化的影响. 甘肃农业大学硕士学位论文: 30–37. |
| [] | 李兆亮, 原永兵, 刘成连, 等. 1998. 水杨酸对叶片抗氧化酶系的调节作用. 植物学报, 40(4): 356–361. |
| [] | 梁建萍, 牛远, 谢敬斯, 等. 2007. 不同海拔华北落叶松针叶三种抗氧化酶活性与光合色素含量. 应用生态学报, 18(7): 1414–1419. |
| [] | 蔺经, 李晓刚, 颜志梅, 等. 2004. 外源水杨酸对山桃幼苗耐冷性的影响. 湖北农学院学报, 24(1): 51–53. |
| [] | 吕军芬, 郁继华. 2004. 水杨酸对西瓜抗冷性生理指标的影响. 甘肃农业大学学报, 39(1): 62–65. |
| [] | 罗毅, 夏国军, 姜玉梅, 等. 2006. 施用有机酸对烤烟生长发育的影响. 安徽农业科学, 34(24): 6524–6526. DOI:10.3969/j.issn.0517-6611.2006.24.060 |
| [] | 束良佐, 李爽. 2002. 水杨酸浸种对水分胁迫下玉米幼苗某些生理过程的影响. 南京农业大学学报, 25(3): 9–11. |
| [] | 宋金凤, 刘永, 崔晓阳. 2004. 毛细管气相色谱法测定森林凋落物中的有机酸. 林业科学, 40(4): 185–188. DOI:10.11707/j.1001-7488.20040433 |
| [] | 宋金凤, 崔晓阳, 王政权. 2008. 柠檬酸/柠檬酸盐对暗棕壤磷的释放效应及机制. 水土保持学报, 22(2): 182–186. |
| [] | 王小华, 庄南生. 2008. 脯氨酸与植物抗寒性的研究进展. 中国农业通报, 24(11): 398–402. |
| [] | 吴楚, 王政权. 2002a. 外源水杨酸对水曲柳幼苗遭受冷害后抗光氧化能力的影响I. 对抗氧化酶活性的影响.湖北农学院学报, 22(4): 303–308. |
| [] | 吴楚, 王政权. 2002b. 冰冻条件下外源SA对水曲柳幼苗叶片内抗氧化酶的影响. 林业科学, 38(5): 54–59. |
| [] | 吴锦程, 陈伟建, 蔡丽琴, 等. 2010. 外源NO对低温胁迫下枇杷幼果抗氧化能力的影响. 林业科学, 46(9): 73–78. DOI:10.11707/j.1001-7488.20100912 |
| [] | 张彩峡. 2010. 低温弱光对紫花苜蓿根颈生理生化指标影响研究. 西北农林科技大学硕士学位论文: 15–29. |
| [] | 张蕊. 2006. 低温下外源水杨酸对水稻幼苗生理生化特性的影响研究. 西南大学硕士学位论文: 22–40. |
| [] | 周艳虹, 喻景权, 钱琼秋, 等. 2003. 低温弱光对黄瓜幼苗生长及抗氧化酶活性的影响. 应用生态学报, 14(6): 921–924. |
| [] | Campos P S, Quartin V, Ramalho J C, et al. 2003. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves of coffea sp.plants. Journal of Plant Physiology, 160(3): 283–293. DOI:10.1078/0176-1617-00833 |
| [] | Dat J F, Lopez-D Delgado H, Foyer C H, et al. 2000. Effects of salicylic acid on oxidative stress and thermotolerance in tobacco. Plant Physiol, 156(5/6): 659–665. |
| [] | Li J Y, Xu R K, Tiwari D, et al. 2006. Mechanism of aluminum release from variable charge soils induced by low-molecular-weight organic acids:Kinetic study. Geochimica et Cosmochimica Acta, 70(11): 2755–2764. DOI:10.1016/j.gca.2006.03.017 |
| [] | Noctor G, Foyer C H. 1998. Ascorbate and glutathione:keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol, 49(1): 249–279. DOI:10.1146/annurev.arplant.49.1.249 |
| [] | Oburger E, Kirk G J D, Wenzel W W, et al. 2009. Interactive effects of organic acids in the rhizosphere. Soil Biology and Biochemistry, 41(3): 449–457. DOI:10.1016/j.soilbio.2008.10.034 |
| [] | Palomo L, Claassen N, Jones D L. 2006. Differential mobilization of P in the maize rhizosphere by citric acid and potassium citrate. Soil Biology and Biochemistry, 38(4): 683–692. DOI:10.1016/j.soilbio.2005.06.019 |
| [] | Pizzeghello D, Zanella A, Carletti P, et al. 2006. Chemical and biological characterization of dissolved organic matter from silver fir and beech forest soils. Chemosphere, 65(2): 190–200. DOI:10.1016/j.chemosphere.2006.03.001 |
| [] | Prasad T K, Anderson M D, Martin B A, et al. 1994. Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide. Plant Cell, 6(1): 65–74. DOI:10.1105/tpc.6.1.65 |
| [] | Reichard P U, Kretzschmar R, Kraemer S M. 2007. Dissolution mechanisms of goethite in the presence of siderophores and organic acids. Geochimica et Cosmochimica Acta, 71(23): 5635–5650. DOI:10.1016/j.gca.2006.12.022 |
| [] | Scandalio J G.1997.Molecular genetics of superoxide dismutases in plants∥Scandalios J G.Oxidative Stress and the molecular biology of antioxidant defenses.Cold Spring Harbor:Cold Spring Harbor Laboratory Press, 545-546. |
| [] | Schmitt A, Glaser B. 2011. Organic matter dynamics in a temperate forest soil following enhanced drying. Soil Biology and Biochemistry, 43(3): 478–489. DOI:10.1016/j.soilbio.2010.09.037 |
| [] | TolráR P, Poschenrieder C, Luppi B, et al. 2005. Aluminium-induced changes in the profiles of both organic acids and phenolic substances underlie Al tolerance in Rumex acetosa L. Environmental and Experimental Botany, 54(3): 231–238. DOI:10.1016/j.envexpbot.2004.07.006 |
| [] | Tsuchidaa H, Kozukue N, Han G-P, et al. 2010. Low-temperature storage of cucumbers induces changes in the organic acid content and in citrate synthase activity. Postharvest Biology and Technology, 58(2): 129–134. DOI:10.1016/j.postharvbio.2010.06.006 |
| [] | Zeng F R, Chen S, Miao Y, et al. 2008. Changes of organic acid exudation and rhizosphere pH in rice plants under chromium stress. Environmental Pollution, 155(2): 284–289. DOI:10.1016/j.envpol.2007.11.019 |