 2012, Vol. 48
2012, Vol. 48文章信息
- 谭小梅, 周志春, 金国庆, 张一
- Tan Xiaomei, Zhou Zhichun, Jin Guoqing, Zhang Yi
- 马尾松二代无性系种子园遗传多样性和交配系统分析
- Genetic Diversity and Mating System Analysis of Pinus massoniana in a Second-Generation Clonal Seed Orchard
- 林业科学, 2012, 48(2): 69-74.
- Scientia Silvae Sinicae, 2012, 48(2): 69-74.
-
文章历史
- 收稿日期:2011-06-29
- 修回日期:2011-10-28
-
作者相关文章



2. 重庆市林业科学研究院 重庆 400036
2. Chongqing Academy of Forestry Chongqing 400036
林木种子园由优树无性系或家系组成,是林木良种繁育的主要形式,也是育种系统的重要组成部分。因此,经营种子园的主要目的是为林业生产提供大量遗传品质优良的种子。然而影响种子品质的因素很多,已有许多研究(Lexer et al., 2000; El-Kassaby et al., 2007)已经表明,交配系统状况是影响种子园种子遗传品质的重要因素。所谓交配系统,是指对生物有性繁殖过程中配子结合类型、模式、影响因素和机制的状态反应,它包含了群体间或群体内个体间基因交流的全部过程,决定着后代群体的基因型分布和种群动态(张大勇等,2001; 何田华等,2001)。因此,了解种子园交配系统的实际情况对于种子园经营与遗传管理有着极其重要的生产实践意义。
影响种子园交配系统因素较多,花期和花量也是其中之一。目前,花量对种子园交配系统的影响主要有2种看法:一种认为,雌球花越多自交率越高,二者成正相关关系(Denti et al., 1988); 另一种认为,偏雌性植株较偏雄性植株个体异交率高(El-Kassaby et al., 1986)。而对于开花物候,开花时间和格局控制着与群体交配系统密切相关的生物和非生物因素在时间上的重叠,从而强烈影响着植物的繁殖成功。已有研究表明,开花物候直接影响授粉效率,其差异不仅存在于无性系间以及同一无性系不同分株之间,同时还存在于同一个体不同树冠部位间(张华新等,2001; 薛康,1992)。因此,研究不同树冠部位(如向阳面和向阴面)以及不同性别取向的植株间的交配系统差异,对于了解其交配系统模式、丰富植物交配系统理论具有重要意义。
马尾松(Pinus massoniana)为我国亚热带东南部典型的针叶树代表树种,也是我国南方具有重要经济和生态价值的主要乡土造林树种(周志春等,2004)。自20世纪80年代初开始营建初级种子园以来,我国有关马尾松种子园的研究取得了显著进展,其中相当一部分研究成果已被广泛应用于生产上,并取得了很好的成果(王章荣等,1990)。然而,在种子园从一代逐步过渡到二代以至更高世代的过程中,建园亲本所受的选择强度逐步加大,亲本间亲缘关系在种子园配置中也难以避免,再加上以往的研究主要是在一代种子园中展开,对国内刚刚进入正常开花结实期马尾松二代种子园的研究还未见报道。因此,本文采用SSR标记对国内较早建立的马尾松二代无性系种子园的交配系统进行研究,并比较亲本与子代种群的遗传多样性,可为马尾松高世代种子园的营建和种子丰产经营提供帮助。
1 材料与方法 1.1 供试材料研究材料来源于浙江省淳安县姥山林场马尾松二代无性系种子园。该园于2002年5月嫁接建成,面积3.7 hm2,共61个二代亲本无性系,分5个生产小区,小区内无性系配置以随机与调整相结合定植,株行距为6 m×8 m,同一无性系分株间距离20 m以上,2008年已进入正常开花结实期。
1.2 样品采集及处理因种子园第5生产小区处于阴坡位置,2008年时开花结实量还很低,故本试验仅在第1~4生产小区内进行,共包括48个无性系。根据已有研究结果(谭小梅等,2011),于2009年在种子园前4个生产小区内,每个小区随机选取1个偏雌型和1个雌雄均衡型无性系单株,采种无性系单株间距离在30 m以上,每个单株在树冠南面和北面中上部各采种10个球果。将采种无性系单株树冠南面和北面球果种子于2010年3月中旬按正常的生产管理方式分别培养子代苗,在幼苗长到10 cm左右时,每个采种单株按树冠南面和北面随机各取20个子代苗嫩叶带回实验室放入-70 ℃冰箱中备用。对种子园亲本采其当年生的幼嫩针叶来确定其基因型。
1.3 总DNA提取DNA的提取采用传统CTAB法(Clark et al., 1998),1%琼脂糖凝胶电泳和紫外检测DNA的浓度与纯度。
1.4 SSR-PCR扩增反应体系10 μL:含1×buffer缓冲液,0.5mmol·L-1的dNTP,0.5 μmol·L-1 SSR引物,30 ng基因组DNA和0.5个单位的Taq酶(TaKaRa生产)。PCR扩增反应程序为: 94 ℃预变性5 min; 94 ℃ 40 s,53 ℃ 30 s,72 ℃ 40 s,35个循环; 最后72 ℃ 10 min。PCR产物混合等体积的加样缓冲液,95 ℃变性5 min后用6%的聚丙烯酰胺变性凝胶电泳分离片段。用银染法显示SSR片段并拍照。
1.5 引物筛选本研究应用的SSR引物是从已经公开发表的文献中得到的(Zhou et al., 2002; 艾畅等,2006; 张薇等,2009; 张冬梅等,2007)。共合成70对SSR引物进行初筛,获得有产物、主带明显的引物49对,复筛后,选出多态性高且稳定的SSR引物12对。位点RPS160的部分样品扩增结果如图 1。

|
图 1 引物RPS160在部分样品中的扩增结果 Fig.1 The part of amplification products of primer RPS160 |
采用人工读带法,根据条带的迁移对其进行判读,采用1,2,3,4,……记录电泳条带数据。排除模糊不清和无法准确标识的带。应用混合交配模型MLTR(Ritland,2008)程序估计单位点异交率、多位点异交率、近交指数及其标准差等。利用软件POPGENE 1.32(Yeh et al., 2000)计算亲、子代的遗传多样性,分析的参数有:等位基因数(A)、有效等位基因数(Ne)、Shannon多样性指数(I)、Nei基因多样度指数(h)、观测杂合度(Ho)、期望杂合度(He)、固定指数(F)。
2 结果与分析 2.1 遗传多样性分析 2.1.1 亲、子代群体的遗传多样性分析利用筛选的12对SSR引物对亲本(61个无性系)和子代群体(320个个体)进行检测(表 1)。在亲本群体中检测到等位基因数2~9个不等,平均为4.167,有效等位基因数为2.223,其中PtTX3116位点的A和Ne最大,分别为9.00和5.787,PR203位点次之。Shannon多样性指数和Nei基因多样度分别为0.850和0.453,稍高于张一等(2010)对马尾松二代育种亲本遗传多样性的研究结果,进一步说明马尾松二代育种亲本确实具备丰富的遗传基础。亲本群体平均期望杂合度(He)和实际观察杂合度(Ho)的均值分别为0.448和0.437,期望值与观察值基本一致。
|
|
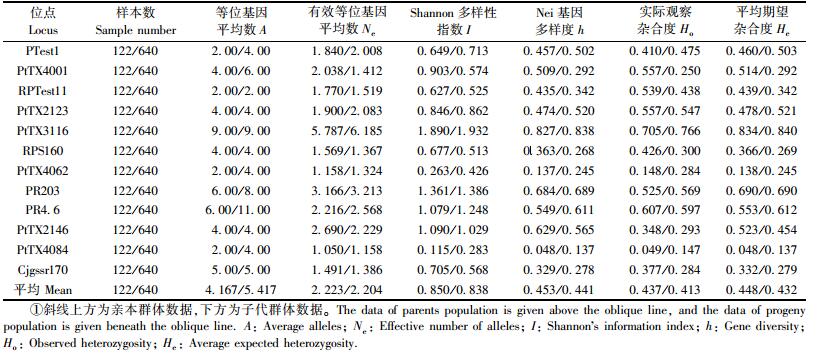
在子代群体中(表 1),12对SSR引物检测到2~11个等位基因,平均为5.417,较亲本群体高出1.25。子代群体的Ne变化在1.158~6.185间,平均为2.204,与亲本群体相当,但仅为其观察等位基因数的一半,其原因是群体中低频等位基因的稳定性不高且对群体遗传变异所起的作用较小。Shannon多样性指数和Nei基因多样度分别为0.838和0.441,均与亲本群体基本保持一致,说明子代能保持亲本所具有的高遗传多样性。12个多态位点的He和Ho平均为0.432和0.413,相对于亲本群体略有减小,说明子代群体基因分布的均匀程度和实际观察到的杂合单株占全部单株比率均较亲本有所降低,但差异不大。据12个位点的Ho和He可计算子代的固定指数F为0.045,存在纯合子过量,但现象不明显。另外,在分析的12个位点中,分别在6个位点检测到2~5个子代特有而亲本没有的等位基因。
2.1.2 树冠南、北面及偏雌和雌雄均衡型植株子代遗传多样性分析对母树树冠南面与北面的子代遗传多样性进行比较(表 2),发现除等位基因数(A)和固定指数(F)外,分析的其余各遗传多样性参数均表现为树冠南面稍高于树冠北面,但差异不显著。说明树冠北面子代遗传多样性并未因其雌、雄球花量较树冠南面减少而有明显的降低。树冠南、北面子代的F值分别为0.031和0.049,均接近于0,不存在明显纯合子过剩现象。
|
|
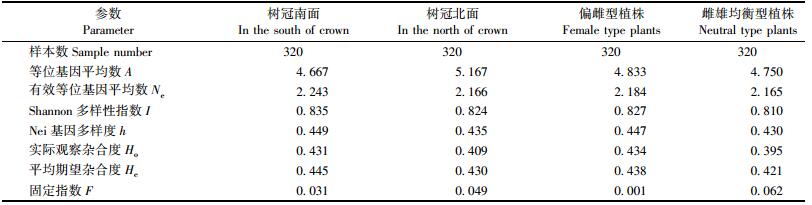
表 2同时对雌雄均衡型和偏雌型植株子代遗传多样性进行了分析。结果表明,遗传多样性参数在2种类型植株子代中基本相当,以偏雌型植株子代略大。偏雌和雌雄均衡型植株的F趋于0,子代基本符合哈温平衡。
2.2 交配系统分析 2.2.1 8个家系的交配系统指标利用MLTR程序对马尾松种子园8个单株的320个子代进行分析(表 3)。由12个位点共同估算出多位点异交率tm=1.098(SD=0.043);单位点异交率ts=1.131(SD=0.046);近交指数(tm-ts)=-0.033(SD=0.047),说明马尾松二代种子园子代亲本的近交现象不显著。对8个家系分别进行交配系统分析(表 3),发现各个家系间的多位点异交率和单位点异交率具有一定的差异。然而何田华等(2001)指出,在实际的估算过程中,偏离混合交配系统模型的一些假设对多位点估算影响较小,而对单位点估算影响较大。因此,研究多注重对多位点异交率的分析。研究的8个家系,多位点异交率变化幅度为1.033 ~1.200,有5个家系的多位点异交率为1.200,其余3个家系的多位点异交率介于1.000~1.200之间。取样家系的近交指数均接近于0,各个家系子代双亲间均不存在明显近交现象。
|
|

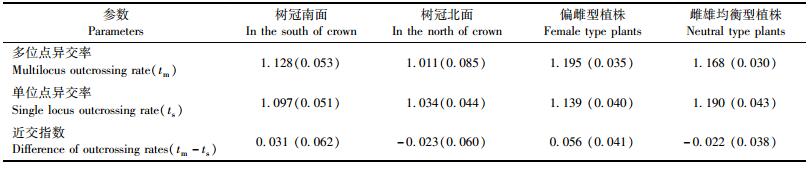
表 4对采种子代进行了树冠方位上异交率的比较。结果表明,树冠南面多位点异交率与单位点异交率均略高于树冠北面,这与上文对树冠南、北面子代遗传多样性的分析结果吻合。对偏雌和雌雄均衡型植株的交配系统指标分析发现(表 4),雌雄均衡型植株与偏雌型植株的多位点异交率基本相当,分别为1.168和1.195,说明马尾松雌雄均衡型植株并未因其雌、雄球花量比例较偏雌型植株减小而呈现异交率明显降低的现象。树冠南面和偏雌型植株的近交指数分别为0.031和0.056,子代双亲存在一定的近交现象,但现象不明显。
|
|
已有研究表明,松属(Pinus)植物是遗传多样性较高的物种,但是不同树种间或同一树种不同群体间的遗传多样性水平是不同的,甚至差异很大。如在松属种子园中,红松(P.koraiensis)的平均期望杂和度(He)为0.601(Feng et al., 2010),海岸松(P.pinaster)为0.784(Fernandes et al., 2008),马尾松一代实生种子园和无性系种子园的He分别在0.529 ~0.575(龚佳等,2007; 张薇等,2008)和0.479 ~0.493(艾畅等,2006; 王鹏良,2006)。本研究马尾松二代无性系种子园子代群体的He为0.432,较前几者有一定程度的降低,分析可能与该种子园刚刚步入正常开花结实期,亲本无性系间雌、雄球花量还未完全稳定有关,也有可能因试验的抽样误差所造成。但是,与资源冷杉(Abies ziyuanensis)的0.337(Tang et al., 2008)、板栗(Castanea mollissima)的0.381(黄武刚等,2009)相比,本研究的马尾松二代无性系种子园子代仍然具有比较丰富的遗传多样性,尤其与马尾松一代无性系种子园相比,其遗传基础没有表现出显著降低的现象。
文中检测到,8个家系间的异交率具有一定的差异。分析这种差异,主要与各个取样亲本同园内其他亲本的亲缘关系远近程度、个体自交可育程度、自身花粉的充裕程度以及所处的位置不同有关。以往的研究表明,松属树种具有典型的高异交率。例如:日本柳杉(Cryptomeria japonica)的异交率为0.944(Moriguchi et al., 2009),红松为0.966(Feng et al., 2011),欧洲赤松(P. sylvestris)为0.950(Korshikov et al., 2010),北美乔松(P. strobus)和扭叶松(P. contorta)分别为1.020和1.000(王崇云等,1999)。本研究获得的马尾松二代种子园多位点异交率为1.098,与松属其他树种一样,具有较高的异交水平,这主要与研究种子园所处的特殊地理环境和马尾松自身的交配机制有关。另外,张薇等(2009)曾对马尾松一代种子园连续2年的异交率进行了调查,分别为1.200和1.072,本文的研究结果与其基本相当,说明马尾松二代种子园仍然保持着一代所具有的高异交水平,并没有表现出很多育种家担心的随建园亲本所受选择强度的增加及亲本间亲缘关系的存在,其子代亲本异交率降低、近交率增加的现象,这从取样时对家系子代生长情况的调查以及子代群体的近交指数(tm-ts=-0.033)接近于0均可看出。本文同时采用固定指数F和近交指数(tm-ts)对种子园的子代群体、母树树冠南北面以及偏雌和雌雄均衡型植株子代近交水平进行了分析,其结果表明,除母树树冠北面和雌雄均衡型植株子代近交指数和固定指数估算值略有差异外,整个子代群体以及树冠南面和偏雌性植株的固定指数和近交指数估计值均基本一致,说明本文研究的马尾松二代无性系种子园子代群体基本符合哈温平衡,子代亲本间不存在明显近交现象。
张薇等(2009)对马尾松种子园母树树冠不同方位的交配系统的研究结果表明,单位点异交率在南面较北面要高,而多位点异交率在南、北两面没有差异。然而本文的研究结果却发现,树冠南面的多位点与单位点异交率均较北面高,可能与2个种子园的坡向和所处地理位置不同有关。分析树冠南面异交率较北面高的原因:其一,与树冠南、北面的雌、雄球花量差异以及所受光照时间不同而造成树冠北面开花物候(雌、雄球花花期)较南面延迟有关(张华新等,2001; 薛康,1992)。因为在种子园花量调查的研究中发现,树冠南面雌、雄球花量普遍多于树冠北面,当雌、雄球花花期相当时,受日光照更充足的树冠南面的雌花更容易接受到更多的外源花粉,其异交率也会随之增大。其二,本文研究的种子园坐西北向东南,位于千岛湖区,周边巨大的水体面积和外部四周的群山环抱形成的独特立体气候和水域气候,使种子园内气流呈回旋模式,加上授粉期内园区主风方向又以东北风为主,因此可能造成树冠南面异交率较北面要高。一般来说,植物在生殖过程中,防止自交的主要机制是自交不亲和,而马尾松属于自交亲和的植物,植株大多为雌雄同株异花,在防止自交衰退的机制上,通常表现为同株上的雄球花较雌球花先开放,即雄球花花期较雌球花提前,这可能就是本研究发现马尾松雌雄均衡型植株异交率并未较偏雌型植株降低的主要原因。然而,因本文仅分别对4个偏雌和4个雌雄均衡型植株进行了单年份分析,因此其准确的相关结论还需后期对更多植株的多年度分析。
已有研究表明,对于风媒授粉树种,种子园中较高的自交率会导致子代群体经济性状的较大分化和平均水平的下降,并且影响种子的生产效率,产生大量空籽(张华新等,2002),因此合理的选择和配置家系或无性系是种子园保持高水平异交率的关键。本试验所得的遗传多样性参数和交配系统指标显示,本研究的马尾松二代种子园子代仍具有丰富的遗传多样性,无性系间的基因交流相对充分,子代亲本近交现象不明显,种子园在营建时家系选择和配置相对比较成功。据此,在营建高世代种子园时,建议可从影响种子园交配系统的众多因子出发,在园址选择、亲本配置、密度管理、花粉管理等方面,尽量减少近交(包括自交)的不良影响,以达到提高高世代种子园产量与品质的双重保证。
| [] | 艾畅, 徐立安, 赖焕林, 等. 2006. 马尾松种子园的遗传多样性与父本分析. 林业科学, 42(11): 146–150. DOI:10.3321/j.issn:1001-7488.2006.11.026 |
| [] | Clark, 顾红雅, 瞿礼嘉, 等. 1998. 植物分子生物学实验手册. 北京: 高等教育出版社; 海德堡: 德国施普林格出版社. |
| [] | 龚佳. 2007. 马尾松实生种子园遗传多样性研究. 南京林业大学硕士学位论文: 21–26. |
| [] | 何田华, 葛颂. 2001. 植物种群交配系统、亲本分析以及基因流动研究. 植物生态学报, 25(2): 144–154. |
| [] | 黄武刚, 程丽莉, 周志军, 等. 2009. 板栗野生居群与栽培品种间叶绿体微卫星遗传差异初探. 林业科学, 45(10): 62–68. DOI:10.11707/j.1001-7488.20091011 |
| [] | 谭小梅, 金国庆, 张一, 等. 2011. 截干矮化马尾松二代无性系种子园开花结实的遗传变异. 东北林业大学学报, 39(4): 39–42. |
| [] | 王崇云, 党承林. 1999. 植物的交配系统及其进化机制与种群适应. 武汉植物学研究, 17(2): 163–172. |
| [] | 王鹏良. 2006. 马尾松无性系种子园多年份子代遗传多样性分析. 南京林业大学硕士学位论文: 17–23. |
| [] | 王章荣, 秦国峰, 陈天华. 1990. 马尾松种子园建立技术论文集. 北京, 学术书刊出版社. |
| [] | 薛康. 1992. 油松开花结实规律的研究. 北京林业大学学报, 14(1): 34–38. |
| [] | 张大勇, 姜新华. 2001. 植物交配系统的进化、资源分配对策与遗传多样性. 植物生态学报, 25(2): 130–143. |
| [] | 张冬梅, 杨娅, 沈熙环, 等. 2007. 油松SSR-PCR引物筛选及反应体系的建立. 北京林业大学学报, 2(9): 13–17. |
| [] | 张华新, 沈熙环. 2002. 林木种子园生殖系统研究进展. 林业科学, 38(2): 129–134. DOI:10.11707/j.1001-7488.20020222 |
| [] | 张华新, 陈丛梅. 2001. 油松无性系开花物候特点的研究. 林业科学研究, 14(3): 288–296. |
| [] | 张薇, 龚佳, 季孔庶. 2008. 马尾松实生种子园遗传多样性分析. 分子植物育种, 4(6): 717–723. |
| [] | 张薇, 龚佳, 季孔庶. 2009. 马尾松实生种子园交配系统分析. 林业科学, 45(6): 22–26. DOI:10.11707/j.1001-7488.20090605 |
| [] | 张一, 谭小梅, 周志春, 等. 2010. 马尾松二代育种亲本主要生长性状和ISSR遗传变异. 分子植物育种, 3(8): 501–510. |
| [] | 周志春, 金国庆, 秦国峰, 等. 2004. 马尾松纸浆材重要经济性状配合力及杂种优势分析. 林业科学, 40(4): 52–57. DOI:10.11707/j.1001-7488.20040409 |
| [] | Denti D, Scheon D J. 1988. Self-fertilization rates in white spruce: effects of pollen and seed production. Heredity, 79(4): 284–288. |
| [] | El-Kassaby Y A, Parkinson J, Davitt W J B. 1986. The effect of crown segment on the mating system in a Douglas-fir(Pseudotsuga menziesii(Mirb)Franco)seed orchard. Silvae Genetica, 35(4): 149–155. |
| [] | El-Kassaby Y A, Stoehr M U, Reid D, et al. 2007. Clonal-row versus random seed orchard designs: interior spruce mating system evaluation. Canadian Journal of Forest Research, 37(3): 690–696. DOI:10.1139/X06-248 |
| [] | Feng F J, Sui X, Chen M M, et al. 2010. Mode of pollen spread in clonal seed orchard of Pinus koraiensis. Journal of Biophysical Chemistry, 1(1): 33–39. DOI:10.4236/jbpc.2010.11004 |
| [] | Feng F J, Zhao D, Sui X, et al. 2011. Study on mating system of Pinus koraiensis in natural population based on cpSSR technology. Advanced Materials Research, 183-185: 700–704. DOI:10.4028/www.scientific.net/AMR.183-185 |
| [] | Fernandes L, Rocheta M, Cordeiro J, et al. 2008. Genetic variation, mating patterns and gene flow in a Pinus pinaster Aiton clonal seed orchard. Ann For Sci, 65(7): 706. DOI:10.1051/forest:2008049 |
| [] | Korshikov I I, Demkovich A E. 2010. Genetic polymorphism of plus-tree clones and their seed progeny in the scotch pine clone plantation. Cytology and Genetics, 44(1): 28–36. DOI:10.3103/S0095452710010068 |
| [] | Lexer C, Heinze B, Gerber S, et al. 2000. Microsatellite analysis of maternal half-sib families of Quercus robur, pedunculate oak: inferring the number of pollen donors from the offspring. Theoretical and Applied Genetics, 100(6): 858–865. DOI:10.1007/s001220051362 |
| [] | Moriguchi Y, Yamazaki Y, Taira H, et al. 2009. Mating patterns in an indoor miniature Cryptomeria japonica seed orchard as revealed by microsatellite markers. New Forests, 39(3): 261–273. |
| [] | Ritland K.2008. Multilocus mating system program MLTR v3.2.http: ∥genetics.forestry.ubc.ca/ritland/programs. https://www.ncbi.nlm.nih.gov/pubmed/18301439 |
| [] | Schmidtling R C, Hipkins V. 1998. Genetic diversity in longleaf pine(Pinus palustris): influence of historical and prehistorical events. Can J For Res, 28(8): 1135–1145. DOI:10.1139/x98-102 |
| [] | Tang S Q, Dai W J, Li M S, et al. 2008. Genetic diversity of relictual and endangered plant Abies ziyuanensis(Pinaceae)revealed by AFLP and SSR markers. Genetica, 133(1): 21–30. DOI:10.1007/s10709-007-9178-x |
| [] | Yeh F C, Yang R.2000.POPGENE v1. 32[ 2000-12].http: ∥www.ualberta.ca. |
| [] | Zhou Y, Bui T, Auckland L D, et al. 2002. Undermethylated DNA as a source of microsatellites from a conifer genome. Genome, 45(1): 91–99. DOI:10.1139/g01-119 |