 2012, Vol. 48
2012, Vol. 48文章信息
- 罗建, 汪书丽, 赵垦田, 杨小林
- Luo Jian, Wang Shuli, Zhao Kentian, Yang Xiaolin
- 拉萨河流域的野生种子植物区系
- Flora of the Seed Plants along the Lhasa River Reaches
- 林业科学, 2012, 48(2): 22-30.
- Scientia Silvae Sinicae, 2012, 48(2): 22-30.
-
文章历史
- 收稿日期:2011-03-16
- 修回日期:2011-06-11
-
作者相关文章



拉萨河流域地处西藏中南部,区内地理位置独特,海拔落差大,地形地貌丰富,自然植被类型多样。由于其特殊的水热环境,为孕育多种植物种类提供了良好的条件。关于西藏植物区系的研究已有一些报道(吴征镒,1987; 李锡文等,1993; 武素功等,1995; 孙航等,2003; 罗建等,2003; 2006; 黄素玲等,2007; 周家福等,2007),这些研究阐明了西藏植物区系概貌以及部分关键、特殊区域植物区系特征。而对于西藏一些河流流域内的植物区系的研究,仅见孙航等(1996; 1997)对喜马拉雅东部雅鲁藏布江大峡湾河谷地区、沈渭寿(1996; 1997)对雅鲁藏布江中游沙地、段代祥等(2005; 2010)对尼洋河流域内河漫滩及巴结湿地和王孙高等(2008)对澜沧江流域内西藏段的研究,对地理位置独特、自然植被类型多样的拉萨河流域植物区系方面的研究尚未见报道。针对这种情况,2007—2010年,笔者对该流域植物的种类及其生态地理分布、植被状况、植物资源等进行较全面的调查,整理出该流域的种子植物名录。在此基础上,对拉萨河流域野生种子植物区系的地理成分进行统计分析,讨论其区系性质、特点及与其他区系的联系,旨在丰富西藏高原植物区系地理的研究资料,为进一步研究当地的植物多样性、植物资源和珍稀植物的开发利用与保护提供参考依据(朱万泽等,2003; 罗建等,2004),为生态环境的保护、建设及管理提供基础资料。
1 研究区概况和研究方法 1.1 研究区概况拉萨河藏语称“吉曲”,流域范围在90°05'—93°20' E,29°20'—31°15' N,发源于念青唐古拉山脉中段北侧的罗布如拉,全长568 km,为雅鲁藏布江最长的一条支流。自源头始向西北流经彭措帕尔和鼓措卧玛两小湖,流入彭措。出彭措后称麦地藏布,至麦曲汇入口以上为上游; 麦曲至桑曲汇入口为中游; 自桑曲以下为下游段,至曲水县附近注入雅鲁藏布江。北部和东北部与怒江流域相邻,东部与帕隆藏布和尼洋河相接,南部为雅鲁藏布江干流,西部和西北部为藏北内流水系。河源地区为平坦湿地,海拔5 150 m,汇入口海拔3 570m,总落差1 580 m,平均坡降2.8‰(中国科学院青藏高原综合科学考察队,1983),其干流段水能蕴藏量1.717×106kW,在雅鲁藏布江各支流中位居第3位。拉萨河流域面积32 471 km2,占雅鲁藏布江流域面积的13.5%,是雅鲁藏布江流域面积最大的一条支流。流域面积占西藏自治区总面积的2.7%,而流域内的人口、耕地却约占全自治区的15%,是西藏工、农、牧业集中的地区。拉萨市就坐落在该河下游。
流域以温带高原季风半干旱河谷以及寒冷半湿润高原气候为主,从下游拉萨、曲水向上游那曲、嘉黎的河源,随着海拔的升高和河谷效应的减弱,气温下降,降水增多。年平均气温-1.9~8.7 ℃,≥10℃的日数50~150天,最暖月平均气温8.9 ~15.5℃,平均最高气温达到15.6~22.6 ℃,年最低温-13.9 ~-2.2 ℃。年降水量400.1~680.6mm,年平均相对湿度45%~61%,年日照时数2 211.8~3 021.7 h(中国科学院青藏高原综合科学考察队,1984),无霜期15~138天。土壤类型以灌丛草原土为主,高山、河谷草甸土和草原土比例也较大,另外还有高山荒漠土、高山寒漠土等多种土壤类型。各种土壤均含石砾较多、透水好、贫瘠。植被类型主要有山地稀疏森林、山地灌丛草原、寒冷半湿润高山草甸、灌丛及流石滩稀疏植被(中国科学院青藏高原综合科学考察队,1988)。
1.2 研究方法依据路线调查、样地调查、野外标本采集、查阅标本馆标本和相关文献资料(吴征镒,1983; 1984; 1986; 1987; 宋永昌,2001),调查路线主要有西、中、东线3条,西线为曲水—堆龙德庆—当雄—那曲; 中线为拉萨—林周—嘉黎; 东线为达孜—墨竹工卡。室内对所有采集的标本进行分类鉴定,根据鉴定结果,加上标本馆查阅历年来在该区采集的标本以及野外调查记录的植物物种,建立拉萨河流域种子植物名录。采用植物区系地理的理论和方法(吴征镒等,1983; 2003a; 2003b; 2004; 2006; 吴征镒,1991; 2003; 王荷生,1992; 李锡文,1996),分析拉萨河流域野生种子植物区系成分及其多样性特征。
2 结果与分析 2.1 植物区系的基本组成拉萨河流域有野生种子植物72科、285属、793种(含变种),种属比为2.78,其中裸子植物3科、3属、6种,被子植物69科、282属、787种。被子植物中双子叶植物有56科、218属、642种,分别占本区野生种子植物科、属、种总数的77.78%,76.49%,80.96%,最为丰富; 其次是单子叶植物,有13科、64属、145种,分别占18.06%,22.46%,18.28%(表 1)。
|
|

拉萨河流域面积仅为西藏总面积的2.7%,从该植物区系与西藏植物区系的比较来看,本区野生种子植物的科、属、种数分别占西藏野生种子植物的43.90%,24.89%,14.97%(吴征镒,1983; 1984; 1985; 1986; 1987),这充分说明拉萨河流域植物物种多样性比较丰富,在西藏植物区系、生物多样性保护中占有较为重要的地位。
2.1.1 种子植物科内属、种的组成在拉萨河流域野生种子植物的72个科中,科内属、种的组成见表 1。含50种以上的科有2个,种数最多者是菊科(30属/105种)(属数/种数,下同)和禾本科(38/75),它们仅占该区总科数的0.28%,所含属数和种数却占总属数的23.86%和总种数的22.70%。菊科、禾本科是世界性分布的几个含数千种以上的大科,在拉萨河流域也有极为丰富的属种,在该植物区系组成中占绝对优势。此外,含20~49种的有蔷薇科(14/44)、毛茛科(13/39)、玄参科(10/38)、豆科(11/38)、石竹科(6/35)、龙胆科(7/33)、虎耳草科(4/31)、十字花科(18/27)、莎草科(7/25)、唇形科(12/21)、罂粟科(4/21)、景天科(4/21)等12个科,这些科中除罂粟科为北温带和南温带间断分布而属温带性质外,全为世界广布科。
以上14科占本区总科数的19.44%,它们的属和种分别占总属、种数的62.46%,69.74%,其包含的属、种数比例较大,具有明显的优势,是该植物区系的重要成分和数量上的优势科,对拉萨河流域植物区系的构建起主导作用。
含19种以下的科共有58个,占总科数80.56%,而所含的属和种数所占比例远比前2类少,这说明本区生境的严酷性,使大多数的科分布到本区的植物种数受到限制(周家福等,2007)。这些科虽然在本区植物区系中物种较少,占从属地位,但在本区的植被构成上仍发挥着重要作用,如小檗科、柏科、马钱科、蓝雪科、麻黄科、荨麻科等,它们同时也表明拉萨河流域植物区系的复杂性和多样性。
2.1.2 种子植物属内种的组成拉萨河流域野生种子植物属内种的组成见表 2。含10种以上的属有13个,占总属数的4.56%;共213种,占总种数的26.86%。20种以上的有虎耳草属(Saxifraga 26种)、龙胆属(Gentiana 22种)、马先蒿属(Pedicularis 21种)、蒿属(Artemisia 21种)4个属。6~10种的属共有20个,占总属数的7.02%;共147种,占总种数的18.54%。主要有蓼属(Polygonum 10种)、黄芪属(Astragalus 10种)、嵩草属(Kobresia 9种)等。2~5种的属有98个,占总属数的34.39%,共279种,占总种数的35.18%。常见的有香薷属(Elsholtzia 5种)、蒲公英属(Taraxacum 5种)、鸢尾属(Iris 5种)、醉鱼草属(Buddleja 4种)、水柏枝属(Myricaria 4)等。仅1种的属有154个,占该区总属数的54.03%,占总种数的19.42%,其中单型属有16个,如独一味属(Lamiophlomis)、绵参属(Eriophyton)、马尿泡属(Przewalskia)、甘松属(Nardostachys)、穗花韭属(Milula)等。由此表明,含1~10种的属在本区系中占有绝对的优势,可见拉萨河流域植物区系中优势属并不明显,本区内植物属的组成是丰富和复杂的。
|
|
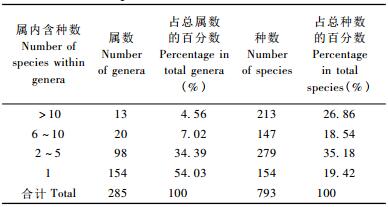
单种属和寡种属极为丰富,二者共占总属数88.42%,所含种数占54.60%。它们是构成拉萨河流域植物区系多元化的主要原因之一,表明拉萨河流域处于较寒冷高山和较温热河谷的某些扩散式分布特征。拉萨河流域的种属比(种/属)2.78,稍高于临近的米拉山(2.39)(罗建等,2003)、西部的阿里西部地区(2.01)(周家福等,2007),但远低于横断山脉(6.0)、青藏高原高寒区(5.27)等地区(李锡文,1995),因此具有属内种系相对贫乏、分化程度较弱的特点。
从上面的统计数据可以看出,在拉萨河流域植物区系中,较大、中型科起到极为重要的作用,而大属的作用并非十分突出。在这些科属内,木本植物较少,多年生草本植物占绝大多数,是本区系的主导物种,这正是以高寒植被类型为主的植物区系所具有的区系特点。
2.2 植物区系的地理成分 2.2.1 科的分布区类型根据关于世界种子植物科的分布区类型系统(李锡文,1996; 吴征镒等,2003a; 吴征镒,2003),拉萨河流域野生种子植物72科可划分为8个分布区类型及变型(表 3)。从科的分布区组成来看,世界分布的科最多,有40科,占总科数的55.56%,主要有菊科、禾本科、蔷薇科、毛茛科、玄参科、伞形科等,包含本地区各种植被类型的建群种和优势种,在种子植物区系中起着重要作用; 其次是温带分布,共18科,占该区非世界科数的56.25%,是拉萨河流域植物区系的主要构成成分,其中北温带和南温带间断分布最多,有柏科、杨柳科、桦木科、罂粟科、胡颓子科等10个科; 北温带分布有忍冬科、松科、杉叶藻科等5科; 欧亚和南美温带间断分布有麻黄科和小檗科; 旧世界温带分布有柽柳科1科,这些科内的种构成林地或灌丛的建群种或优势种,同时表现该区系组成中科的温带性质。热带分布有14科,占该区非世界科数的43.75%,其中泛热带分布最多,包括荨麻科、蒺藜科、大戟科、卫矛科、锦葵科等9个科; 另外热带亚洲和热带美洲间断分布有紫茉莉科、苦苣苔科、川续断科3科; 热带亚洲-热带非洲-热带美洲(南美洲)有马钱科和鸢尾科2科等,表明拉萨河流域种子植物区系在发生发展过程中曾经历过与热带相联系的历史渊源。在本区未见中国特有分布科。
|
|

从科的分布区类型可以看出,拉萨河流域植物科的地理成分主要以世界分布为主,它们大多是主产在温带地区的世界性大科,如菊科、禾本科、龙胆科、莎草科等,仍可以看出该区温带为主导的性质。而且这些世界科所分布到本区的属中,多数属和种属于以温带分布为主的属和种。从表 3可以看到,世界广布科有40个之多,而种只有13个,热带性质科有14个,种只有10个,比例极小。
2.2.2 属的分布区类型中国种子植物属的15个分布区类型、31个变型中(吴征镒,1991; 吴征镒等,2003b; 2004),拉萨河流域有15个类型及其11个变型(表 3),反映其种子植物区系联系的广泛性和复杂性。
世界分布49属,占总属数的17.19%,以草本和灌木为主,如早熟禾属(Poa)、藜属(Chenopodium)、茄属(Solanum)、沟酸浆属(Mimulus)、酸模属(Rumex)、眼子菜属(Potamogeton)等,其中很多几乎只见于路边、荒坡、草丛、水塘,在森林群落中并不多见。
热带分布的6种类型(类型2~7)在拉萨河流域都存在,共计31属,占该区非世界属数的13.14%。其中: 1)泛热带分布及其变型最多,计24属,占该区非世界属数的10.17%,其中很多是河谷和石质山坡的常见成分,如麻黄属(Ephedra)、蒺藜属(Tribulus)、醉鱼草属(Buddleja)、草沙蚕属(Tripogon)、须芒草属(Andropogon)、虎尾草属(Chloris)、曼陀罗属(Datura)、球柱草属(Bulbostylis)等,该类型的属在拉萨河流域内分布种数较少。2)旧世界热带有香茅属(Cymbopogon)和天门冬属(Asparagus)2个,而后者的种类已接近旧世界温带; 3)热带亚洲至热带非洲及变型香茶菜属(Rabdosia)和紫金标属(蓝雪花属)(Ceratostigma)2个; 4)热带亚洲和热带美洲间断分布紫茉莉属(Mirabilis)1个; 5)热带亚洲至热带大洋洲阔蕊兰属(Peristylus)1个; 6)热带亚洲(即热带东南亚至印度-马来,太平洋诸岛)有苦荬菜属(Ixeris)1人。
本区的热带成分比例较小,主要分布在西南部分河谷地区,表明热带分布属在拉萨河流域种子植物区系组成中处于从属地位,其影响十分有限但较有意义。就这仅有的31个属的现代分布区范围来看,它们不仅都延伸到温带地区,并且其中一些种的主要分布区是在温带地区,这说明本区系虽与热带植物区系有联系,但这种联系的程度已微乎其微。特别是本区干旱、多风、寒冷、高辐射的生态环境,已难适宜更多的热带成分分布于此。虽然也有一些属可勉强分布于此,但属中多由少数种(5种以下)作为其代表,且这些种已经随着这里的环境相当特化,这些热带成分植物主要在本区特殊的区域—干旱、温暖的河谷地段分布,反映本区处于热带植物分布区的北部边缘。
温带分布(类型8~11,14)共有187属,占该区非世界属数的79.24%,在拉萨河流域植物区系中居绝对优势,是该流域种子植物区系的主要地理成分。其中: 1)温带性质的属中,数量最多的北温带分布型及其3个变型有106属,占该区非世界分布属的44.92%,占本区温带属的56.68%。2)居第2位的是旧世界温带分布及其变型共有41属,占该区非世界属数的17.37%和占本区温带属21.93%。3)东亚(东喜马拉雅-日本)及其变型有27属,占该区非世界分布属的11.44%。东亚是被子植物早期分化的一个关键地区(路安民,1999),其中最突出的便是中国-喜马拉雅成分变型成分23属,占东亚属的85.19%,如丛菔属(SolmsLaubachia)、矮泽芹属(Chamaesium)、蓝钟花属(Cyananthus)、穗花韭属(Milula)等。它们是本区高寒草甸和高寒灌丛的伴生植物,也有少数高山流石坡种类,它们的出现反映其适应高原、高山寒旱环境的分布特点,也表明本区生态地理特征和严寒的生态环境对东亚类型成分的选择及其分布范围的影响,这些属的存在反映出该区与东亚,尤其与东喜马拉雅许多重要植物类群有一定关系,是高山植物的现代分布分化中心,甚至可能是其中一些属下等级的起源中心(Wu,1988; 孙航,2002)。4)温带亚洲分布,在拉萨河流域有8个属,多数仅有1~2种,本类型在本区植被组成上的作用较小。其中狗娃花属(Heteropappus)在中下游宽谷地段比较常见; 锦鸡儿属(Caragana)在高山稀疏灌丛草甸中构成灌丛植被的建群成分或优势成分; 细柄茅属(Ptilagrostis)和亚菊属(Ajania)分别衍生于北温带大属—针茅属(Stipa)和菊蒿属(Tanacetum),在拉萨河流域有广泛分布,这不仅表明拉萨河流域植物区系的年轻和衍生的性质,而且还表明以高寒因子为主的高山特化现象在本区的普遍存在(周家福等,2007)。5)东亚和北美洲间断及其变型有5属,如黄华属(Thermopsis)、燕麦属(Avena)等,反映该区与东亚和北美洲在区系上有一定关系。此外如寒原荠属(Aphragmus)这样经过长期衍化的属,已有明显的高山特化特征,在该区的分布种类亦多以中国-喜马拉雅类型为主。
中亚分布和地中海区、西亚至中亚分布及其变型,虽然主体分布在新疆等干旱地区(Wu,1988),但仍可以在拉萨河流域找到。前者在拉萨河流域有10属分布,全部为中亚东部至喜马拉雅和中国西南分布变型,如藏芥属(Phaeonychium); 后者有群心菜属(Brassica)1个,反映干旱环境对拉萨河流域植物区系产生深远的影响,并且在这些地区高山植物区系的形成中扮演较为重要的角色。
现存的许多中国-喜马拉雅成分是古地中海的后裔,汤彦承等(1996)指出特提斯第三纪植物群是东亚被子植物区系的一个重要源头,一些中国-喜马拉雅成分的特征属如刺参属(Oplopanax)、独一味属(Lamiophlomis)、绵参属(Eriophyton)、扭连钱属(Phyllophyton)等是古地中海退却后,随着喜马拉雅山脉隆升为陆地,适应高山荒漠而由其地中海的祖先类群分化、衍生出来的年轻类群(吴征镒等,1982; 李锡文,1989)。这些类群在拉萨河流域的存在也反映本植物区系有着深远的古地中海渊源。
中国特有属分布有7个,占该区非世界属数的2.97%,除合头菊属(Syncalathium)外,几乎全为单种属,其中被菊属(Chlamydites)为西藏特有,马蹄黄属(Spenceria)、舟瓣芹属(Sinolimprichtia)、环根芹属(Cyclorhiza)、合头菊属分布主产于西南,马尿泡属(Przewalskia)产于唐古特地区,三蕊草属(Sinochasea)产于唐古特-喜马拉雅地区,多是高原特化性质的属种,这证实本植物区系的年轻性。
2.2.3 种的分布区类型拉萨河流域共有野生种子植物793种(吴征镒,1983; 1984; 1985; 1986; 1987),可归入除热带亚洲和热带美洲间断分布、热带亚洲至热带大洋洲分布以及东亚和北美洲间断类型以外的12个分布区类型及其15个变型(表 3)。
世界分布种共有13种,全部为草本植物,主要是田间杂草、水生植物或伴人植物。常见的有曼陀罗(Datura stramonium)、牛膝菊(Galinsogaparviflora)、金鱼藻(Ceratophyllum demersum)、高山梯牧草(Phleum alpinum)等。
热带分布类型(类型2~7)共计10种,占总种数的1.28%,在拉萨河流域区系和植被组成中不具有重要作用。全部为草本植物。常见的有稗(Echinochloa crusgali)、星鳞狸藻(Utricularia stellaris)、喜马拉雅草沙蚕(Tripogon hookerianus)、尼泊尔酸模(Rumex nepalensis)等。
温带分布类型(类型8~11,14)共有381种,占总种数的48.85%。北温带分布及其变型共有56种,占总种数的7.18%,草本植物占绝对的优势,如紫羊茅(Festuca rubra)、珠芽蓼(Polygonumviviparum)等在亚高山草甸成为建群种或优势种,以及许多水生植物,如红线草(Potamogeton pectinatus)、水葱(Scirpus validus)、浮萍(Lemnaminor)等。常见的还有毛蕊花(Verbascum thapsus)、萹蓄(Polygonum aviculare)、蕨麻(Potentilla anserina)、广布野豌豆(Vicia cracca)等。
旧世界温带分布及其变型共有42种,占总种数的5.38%,在拉萨河流域植物区系组成中占有一定的地位,其中绝大多数是草本植物,有少数乔灌木。常见的灌木有白柳(Salix alba)、小叶金露梅(Potentilla parvifolia)、小叶栒子(Cotoneaster microphyllus)等。常见草本植物有印度草木犀(Melilotus indica)、紫苜蓿(Medicago sativa)、野燕麦(Avena fatua)、大车前(Plantago major)、天蓝苜蓿(Medicago lupulina)等。
温带亚洲分布共有25种,占3.21%,常见的乔木有白桦(Betula platyphylla),是落叶阔叶林的建群种或优势种。单子麻黄(Ephedra monosperma)常在干旱山坡形成斑块状低矮灌丛群落,木本植物还有高山绣线菊(Spiraea alpina)。草本植物常见的有垂穗披碱草(Elymus nutans)、毛叶老牛筋(Arenariacapillaris)、鼠麴草(Gnaphalium affine)、沙蒿(Artemisia desertorum)等。
东亚分布共有258种,占总种数的33.08%,占温带分布的67.72%,其中中国-喜马拉雅分布变型253种,占东亚分布的98.06%,在拉萨河流域植物区系组成中占有绝对的优势,其中许多是不同植被中的优势种或特征种。常见的木本植物有香柏(Sabina pingii var. wilsonii)、藏麻黄(Ephedrasaxatilis)、山岭麻黄(Ephedra gerardiana)、硬叶柳(Salix sclerophylla)、有棱小檗(Berberis angulosa)、雪层杜鹃(Rhododendron nivale)、岩生忍冬(Lonicerarupicola)、匍匐栒子(Cotoneaster adpressus)、小苞水柏枝(Myricaria wardii)等。草本植物种类所占比例很大,常见的有圆穗蓼(Polygonum macrophyllum)、花葶驴蹄草(Caltha scaposa)、蓝翠雀花(Delphiniumcaeruleum)、西藏铁线莲(Clematis tenuifolia)、钉柱委陵菜(Potentilla saundersiana)、劲直黄耆(Astragalusstrictus)、斑唇马先蒿(Pedicularis longiflora var.tubiformis)、拉萨狗娃花(Heteropappus gouldii)、毛香火绒草(Leontopodium stracheyi)、高山嵩草(Kobresiapygmaea)、穗花韭(Milula spicata)等,它们多为草本层或草本群落的优势种或建群种。
地中海区、中亚分布(类型12~13)有65种,占总种数的8.33%,分别是鬼箭锦鸡儿(Caraganajubata)、长刺茶藨子(Ribes alpestre)、独行菜(Lepidium apetalum)、白草(Pennisetum centrasiaticum)、天山千里光(Senecio tianshanicus)、卵萼花锚(Halenia elliptica)、喉毛花(Comastoma pulmonarium)、展苞灯心草(Juncus thomsonii)、细果角茴香(Hypecoum leptocarpum)、披针叶黄华(Thermopsis lanceolata)和水葫芦苗(Halerpestes cymbalaria)。这一类型反映拉萨河流域植物区系与地中海、西亚和中亚较为密切的联系。
中国特有分布共有324种,占总种数的41.54%,在拉萨河流域植物区系组成中占有绝对的优势。根据其地理分布特点划分如下。
1) 西藏特有135种,其中拉萨河流域特有37种,如砂生小檗(Berberis sabulicola)、多刺锦鸡儿(Caragana spinifera)、拉萨虫实(Corispermum lhasaense)、古临无心菜(Arenaria littledalei)、耐国蝇子草(Silene zhoui)、伏毛直序乌头(Aconitum richardsonianum var. pseudosessiliflorum)、拉萨厚棱芹(Pachypleurum lhasanum)、拉萨龙胆(Gentianawaltonii var. lhasaensis)、褶苞香青(Anaphalisplicata)、喙马唐(Digitaria cruciata var.esculenta)等。
2) 西南特有102种,如大果圆柏(Sabinatibetica)、藏川杨(Populus szechuanica var. tibetica)、小叶野丁香(Leptodermis microphylla)、条裂银莲花(Anemone trullifolia var. linearis)、马蹄黄(Spenceriaramalana)、全萼龙胆(Gentiana lhassica)、草柏枝(Phtheirospermum tenuisectum)、川西小黄菊(Pyrethrum tatsienense)、须芒草(Andropogon yunnanensis)等。
3) 西南、西北(指唐古特)特有71种,如山生柳(Salix oritrepha)、毛叶绣线菊(Spiraea mollifolia)、窄叶鲜卑花(Sibiraea angustata)、矮锦鸡儿(Caragana maximovicziana)、马尿泡(Przewalskia tangutica)、山莨菪(Anisodus tanguticus)、蓝玉簪龙胆(Gentiana veitchiorum)、中华紫茉莉(Mirabilis himalaica var.chinensis)、铁棒锤(Aconitum pendulum)、毛果婆婆纳(Veronica eriogyne)、丝颖针茅(Stipa capillacea)等。
4) 西南、华中5种,有川滇绣线菊(Spiraea schneideriana)、峨眉蔷薇(Rosa omeiensis)等。
5) 西南、华北、西北10种,常见者有乌柳(Salix cheilophila)、鹤虱(Lappula myosotis)、秃疮花(Dicranostigma leptopodum)等。
6) 西南、华东(台湾)1种,见有大花三毛草(Trisetum spicatum subsp. alaskanum)。综上所述,可见拉萨河流域特有种类相当丰富,种级水平的特有现象明显,这充分证实本区系植物的年青性及其较强的演化、特化性质。
3 结论与讨论根据以上结果,拉萨河流域野生种子植物区系具有以下特征。
1) 野生种子植物具较高的丰富性与多样性。拉萨河流域内现有野生种子植物72科、285属、793种(含变种),分别占西藏种子植物的43.90%,24.89%,14.97%。属内种系相对贫乏,种属比2.78,分化程度较弱,单种属和寡种属极为丰富。
2) 含20种以上的14个科,包含的属、种数比例较大,具有明显的优势,是该植物区系的重要成分和数量上的优势科,对拉萨河流域植物区系的构建起主导作用。就属的组成来看,单种属和寡种属极为丰富,是构成拉萨河流域植物区系多元化的主要原因之一,说明本区系中属的分化程度较高。
3) 种子植物的区系成分复杂,其中温带成分分布占绝对优势:温带科18个,占总科数的56.25%,温带属194个,占总属数的82.20%,温带种705个,占总种数的90.38%,说明本区系具有明显的温带性质。而热带成分仅在科级分布型种表现出较大的比例,属种级热带成分很少,表明本区系在发生发展过程中曾经历过与热带相联系的历史渊源。
4) 表现出一定程度的特有性,没有中国特有科,中国特有属分布7个,中国特有分布种324个,证实本区系植物的年青性及其较强的演化、特化性质。
5) 拉萨河流域植物区系与中国-喜马拉雅植物区系的联系紧密,同时与地中海区、西亚至中亚植物区系亦有深远的渊源。
由于喜马拉雅强烈隆起的影响,拉萨河流域古地理环境变迁很大,近期自然环境形成的历史短暂(第三纪末开始),在年轻的高山环境中产生一些新的区系成分,大多为喜马拉雅所独有。同时,受半干旱半湿润高原季风气候影响,生态条件严酷,限制了植物区系的发生和发展(中国科学院青藏高原综合科学中国科学院青藏高原综合科学考察队,1982)。植物的分布在流域内,随着海拔升高、气候条件相应变化,由森林、灌丛、草原、草甸至高山流石滩荒漠而变化,区系成分的丰富程度总体上亦随之减小。拉萨河流域地史上受第四纪冰期恶劣气候影响较小,为植物提供良好的生存环境。这种特殊的地质历史和地理环境,对植物区系的发生和发展带来深刻影响。而拉萨河流域植物的热带成分在科级分布型种表现出较大的比例(14科,占总科数的19.44%),保留热带区系的历史痕迹,表现出本区系与热带相联系的历史渊源(第三纪古热带区系),但属种级热带成分很少,反映现代气候对区域内植被组成、发生、发展、迁移及演化的影响。在持续隆升的过程中,复杂的山地地形和逐渐深切的河谷也创造或提供不少的天然避难所,因而形成以温带区系成分为主、多种区系成分并存、耐寒和喜暖成分五方杂处、种类组成丰富、物种的水平替代和垂直分布变化十分显著的植物区系特征。
| [] | 段代祥, 陈贻竹, 赵南先, 等. 2005. 西藏巴结湿地自然保护区种子植物区系的研究. 华南农业大学学报, 26(2): 81–85. DOI:10.7671/j.issn.1001-411X.2005.02.021 |
| [] | 段代祥, 赵南先, 吴兴. 2010. 西藏尼洋河河漫滩种子植物区系研究. 浙江林学院学报, 27(3): 354–359. |
| [] | 黄素玲, 王翠民, 盖连玉. 2007. 藏西草地植物区系特征研究. 内蒙古草业, 19(2): 32–38. |
| [] | 李锡文. 1989. 横断山区唇形科植物的地理分布. 植物研究, 9(2): 103–122. |
| [] | 李锡文. 1995. 云南热带种子植物区系. 云南植物研究, 17(2): 115–128. |
| [] | 李锡文. 1996. 中国种子植物区系统计分析. 云南植物研究, 18(4): 363–384. |
| [] | 李锡文, 李捷. 1993. 横断山脉地区种子植物区系的初步研究. 云南植物研究, 15(3): 217–231. |
| [] | 路安民. 1999. 种子植物科属地理. 北京, 科学出版社: 1-5. |
| [] | 罗建, 边巴多吉, 郑维列. 2003. 西藏米拉山区种子植物区系研究. 南京林业大学学报:自然科学版, 27(6): 18–22. |
| [] | 罗建, 郑维列, 潘刚, 等. 2006. 色季拉山区高山寒带种子植物区系研究. 武汉植物学研究, 24(3): 215–219. |
| [] | 罗建, 郑维列, 邢震, 等. 2004. 西藏米拉山区植物资源特征分析. 浙江林学院学报, 21(3): 275–280. |
| [] | 沈渭寿. 1996. 雅鲁藏布江中游沙地植物区系特征. 植物分类学报, 34(3): 276–281. |
| [] | 沈渭寿. 1997. 雅鲁藏布江中游沙地植物区系与阿拉善荒漠植物区系的比较. 中国沙漠, 17(1): 15–19. |
| [] | 宋永昌. 2001. 植被生态学. 上海, 华东师范大学出版社: 296-562. |
| [] | 孙航, 李志敏. 2003. 古地中海植物区系在青藏高原隆起后的演变和发展. 地球科学进展, 18(6): 852–862. |
| [] | 孙航, 周浙昆. 1996. 喜马拉雅东部雅鲁藏布江大峡湾河谷地区植物区系的特点及来源. 云南植物研究, 18(2): 185–204. |
| [] | 孙航, 周浙昆. 1997. 喜马拉雅东部雅鲁藏布江大峡湾河谷地区种子植物区系的性质和近缘关系. 应用与环境生物学报, 3(2): 184–190. |
| [] | 孙航. 2002. 古地中海退却与喜马拉雅-横断山的隆起在中国喜马拉雅成分及高山植物区系的形成与发展上的意义. 云南植物研究, 24(3): 273–288. |
| [] | 汤彦承, 李良千. 1996. 试论东亚被子植物区系的历史成分和第三纪源头:基于省沽油科、刺参科和忍冬科植物地理的研究. 植物分类学报, 34(5): 453–478. |
| [] | 王荷生. 1992. 植物区系地理. 北京, 科学出版社: 1-180. |
| [] | 王孙高, 袁睿佳, 王宝荣, 等. 2008. 澜沧江(西藏段)流域种子植物区系研究. 云南大学学报:自然科学版, 30(S2): 377–383. |
| [] | 武素功, 杨永平, 费勇. 1995. 青藏高原高寒地区种子植物区系的研究. 云南植物研究, 17(3): 233–250. |
| [] | 吴征镒, 陈心启. 2004. 中国植物志(1卷). 北京:科学出版社: 1–583. |
| [] | 吴征镒, 李锡文. 1982. 论唇形科的进化与分布. 云南植物研究, 4(2): 97–118. |
| [] | 吴征镒, 路安民, 汤彦承, 等. 2003b. 中国被子植物科属综论. 北京, 科学出版社. |
| [] | 吴征镒, 王荷生. 1983. 中国自然地理:植物地理(上册). 北京, 科学出版社: 1-125. |
| [] | 吴征镒, 周浙昆, 李德铢, 等. 2003a. 世界种子植物科的分布区类型系统. 云南植物研究, 25(3): 245–257. |
| [] | 吴征镒, 周浙昆, 孙航, 等. 2006. 种子植物的分布区类型及其起源和分化. 昆明, 云南科技出版社: 1-566. |
| [] | 吴征镒. 2003. 世界种子植物科的分布区类型系统的修订. 云南植物研究, 25(5): 535–538. |
| [] | 吴征镒. 1991. 中国种子植物属的分布区类型. 云南植物研究, 13(z1): 1–139. |
| [] | 吴征镒. 1983. 西藏植物志(1卷). 北京, 科学出版社. |
| [] | 吴征镒. 1984. 西藏植物志(2卷). 北京, 科学出版社. |
| [] | 吴征镒. 1985. 西藏植物志(3卷). 北京, 科学出版社. |
| [] | 吴征镒. 1986. 西藏植物志(4卷). 北京, 科学出版社. |
| [] | 吴征镒. 1987. 西藏植物志(5卷). 北京, 科学出版社. |
| [] | 中国科学院青藏高原综合科学考察队, 1988. 西藏植被, 北京: 科学出版社, 234-251. |
| [] | 中国科学院青藏高原综合科学考察队, 1983. 西藏地貌, 北京: 科学出版社, 56-57. |
| [] | 中国科学院青藏高原综合科学考察队, 1984. 西藏气候, 北京: 科学出版社, 142-170. |
| [] | 中国科学院青藏高原综合科学考察队, 1982. 西藏自然地理, 北京: 科学出版社, 146-160. |
| [] | 周家福, 张锦华, 刘淑珍, 等. 2007. 西藏阿里西部地区种子植物区系研究. 山地学报, 25(5): 608–615. |
| [] | 朱万泽, 范建容. 2003. 西藏珍稀濒危植物区系特征及其保护. 山地学报, 21(z1): 31–39. DOI:10.3969/j.issn.1008-2786.2003.z1.007 |
| [] | Wu Z Y. 1988. Hengduan mountain flora and her significance. Journal of Japanese Botany, 63(9): 297–311. |