 2012, Vol. 48
2012, Vol. 48文章信息
- 周先容, 余岩, 周颂东, 何兴金
- Zhou Xianrong, Yu Yan, Zhou Songdong, He Xingjin
- 巴山榧树地理分布格局及潜在分布区
- Geographic Distribution and Potential Distribution of Torreya fargesii
- 林业科学, 2012, 48(2): 1-8.
- Scientia Silvae Sinicae, 2012, 48(2): 1-8.
-
文章历史
- 收稿日期:2011-01-29
- 修回日期:2012-01-04
-
作者相关文章



2. 长江师范学院生命科学与技术学院 涪陵 408100
2. College of Life Science and Technology, Yangtze Normal University Fuling 408100
巴山榧树(Torreya fargesii)隶属于红豆杉科(Taxaceae)榧树属(Torreya), 为我国特有植物, 收录在《中国物种红色名录》中, 其濒危等级和标准为VU A2c(汪松等, 2004)。榧树属植物共7种, 分布在中国、日本、美国的东南部和西部, 我国有5种, 包括日本榧树(T.nucifera)、榧树(T.grandis)、巴山榧树、长叶榧树(T.jackii)和四川榧(T.parvifolia), 其中日本榧树为引进栽培种, 另外4种均为我国特有种。此外, 本属还有2变种:九龙山榧树(T.grandis var.jiulongshanensis)和云南榧(T.fargesii var.yunnanensis)(康宁等, 1995; Fu et al., 1999; 易同培等, 2006)。根据已有的化石记录, 榧树属始见于中侏罗纪至新第三纪, 在欧洲、亚洲及北美洲均有分布。第四纪冰期后, 在欧洲绝灭, 形成了现代的东亚和北美间断分布, 以我国种类最多, 是一类古老珍稀的孑遗植物, 具有较高的经济价值和学术研究价值(康宁等, 1995)。我国所有榧树属种类均被列为国家Ⅱ级重点保护野生植物(于永福, 1999)。
巴山榧树分布于我国陕西南部, 湖北西部, 四川东部、东北部和西部, 海拔1 000~1 800 m, 散生于林分中。其木材坚硬、结构细致, 可作家具及农具等, 种子可榨油, 是一种非常重要的林木资源(郑万钧等, 1978)。目前国内外对该物种的研究很少, 除作为试验材料用于探讨红豆杉科及榧树属的系统发育外, 仅涉及植物化学成分和形态学研究(席以珍, 1986; 罗士德等, 1994; 康宁等, 1995; 苏应娟, 1997; 陈振德等, 1998; 2000; 汪小全等, 2000; Li et al., 2001)。虽然《中国植物志》(郑万钧等, 1978)、《Flora of China》(Fu et al., 1999)、《四川植物志》(管中天, 1983)和《湖北植物志》(傅书遐等, 2001)等文献对巴山榧树的分布有记录, 但对该物种的地理分布格局及保护生物学等方面的研究还十分缺乏。
物种的分布格局是物种重要的空间特征, 对研究物种的起源、散布及演化有着重要意义, 受到宏观生物学领域众多学者的关注, 成为生物多样性科学的一个基本概念和研究对象, 无论在理论研究还是在制定保护政策上都有重要意义(张清华等, 2000; Bell, 2001; Maurer, 2002; Nee, 2002; 张文驹等, 2003; 郭泉水等, 2005; Zhang et al., 2008)。基于GIS技术的物种分布格局研究已用来预测物种的潜在分布区, 指导物种的保护和资源调查(Funk et al., 1999; Attorre et al., 2007; Beaumont et al., 2007; 徐晓婷等, 2008)。GARP(genetic algorithm for rule-set production)生态位模型是一种基于检验假设的分布预测模型, 通过生态因子和物种分布的反复建模迭代来探索物种分布区的环境特征与研究区域的非随机关系, 能够直观地体现出各种生态因子与物种的关系, 是目前应用最广泛的物种潜在分布区预测方法之一(Adjemian et al., 2006)。本研究以巴山榧树的分布数据为基础, 通过Desktop Garp软件包对其潜在分布区进行GARP模型预测, 同时结合野外调查资料分析讨论该物种的地理分布、种群现状、濒危原因及保护策略等, 为巴山榧树的物种保护、资源普查和迁地保存等提供理论依据。
1 研究方法 1.1 巴山榧树分布数据的收集与处理查询中国数字植物标本馆(http:∥www.cvh.org.cn/)、国家科技部教学标本资源共享平台(http:∥mnh.scu.edu.cn/)数据库, 并实地查阅标本馆中巴山榧树的标本, 共获得284份巴山榧树标本信息。查阅《中国植物志》(郑万钧等, 1978)、《Flora of China》(Fu et al., 1999)、《四川植物志》(管中天, 1983)、《湖北植物志》(傅书遐等, 2001)、《湖南植物志》(刘克明等, 2000)和《河南植物志》(丁宝章等, 1981)等地方植物志及相关研究文献对标本的鉴定和分布信息进行核对, 仅选取其野生分布的信息。在此基础上, 从巴山榧树地理分布数据中选取有确切分布地点的地区作为野外调查目的地, 每个省级行政单位至少1个, 2009—2010年对巴山榧树的分布区进行野外调查。结合标本数据和野外调查数据, 最终获取不重复的分布点57个。所有分布地信息至少精确至县一级, 经纬度分辨率小于0.1°。在物种分布信息收集的基础上, 用ArcGIS软件绘制巴山榧树的地理分布图, 分别以海拔图层和植被覆盖图层作为底图(http:∥biogeo.berkeley.edu/bgm/gdata.php), 精度为30″。
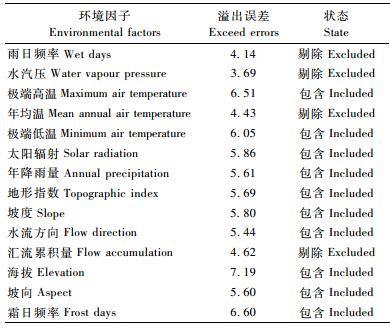
1.2 环境数据的获取GARP所用环境数据集包括14个环境因子, 其中极端高温、极端低温、年均温、年降雨量、雨日频率、水汽压、霜日频率和海拔来自Berkeley大学Worldclim下载中心(http:∥www.worldclim.org/), 所有层数据的投影格式均转换为GCS_WGS_1984。Worldclim数据库对1950—2000年来自世界各地气象站的气候信息采用插值法生成全球气候数据, 空间分辨率可达到30″。太阳辐射、地形指数、坡度、水流方向、汇流累积量和坡向采用软件自带的亚洲数据集(Institute of Gazetteer Research, 1994), 以地理栅格为单位计算。最终预测结果使用从国家基础地理信息系统(http:∥nfgis.nsdi.gov.cn/)下载的1: 400万的中国行政区划图作为分析底图。
1.3 巴山榧树的生态位模型预测在使用Desktop Garp时, 每次运算随机选取50%的数据用于建立模型, 剩下50%的数据用于验证模型误差。创建模型规则时反复运算1 000次或者所有分布点都符合模型规则而终止。在选取最优的环境因子时, 首先将14个环境因子一起进行1 000次模型运算, 将所获得的溢出误差(exceed errors, Desktop Garp结果中的commission项)95%置信区间的算术平均值作为基础溢出误差; 然后使用刀切法分析环境因子对结果的影响(王瑞, 2006), 去除会增大误差的环境因子; 在最终分析中, 重复创建1 000次模型, 并生成ARC格式的图层。运算完毕后, 选取所有内部遗漏误差和外部遗漏误差为零的预测中溢出误差最小的100个模型, 即为最优模型。利用ArcGIS的空间扩展模块, 将最优模型的预测结果进行等权重叠加。叠加后, 每一个栅格的数值为0~100, 这个数值称为重叠系数(overlay value, OV), 代表 100个预测模型中该栅格适合生长巴山榧树的模型数量。OV值越大, 表示该栅格的环境参数越适合巴山榧树生存。根据重叠系数的大小可以将预测的区域划分为:良好生长区(OV ≥ 90%)、可生长区(70% ≤ OV<90%)、过渡区(50% ≤OV<70%)和不适生长区(OV<50%)。
2 结果与分析 2.1 地理分布以海拔图层为底图绘制的巴山地理分布图(图 1)表明, 巴山榧树分布在我国中亚热带至北亚热带的中低山地区(102°35'—116°21' E, 28°51'—34°26' N), 集中分布于秦岭、大巴山、邛崃山、大娄山、巫山、武陵山和大别山, 跨四川、贵州、陕西、甘肃、湖北、湖南、河南、安徽和重庆9省市。标本记录和野外调查显示, 巴山榧树分布在海拔800~2 700 m, 最低处位于湖南省石门县和陕西省旬阳县, 最高处位于四川省宝兴县。巴山榧树分布区虽跨越9省市, 但分布区域狭窄, 呈斑块状分布, 实际面积不大(图 1)。野外调查(表 1)显示, 26个巴山榧树种群绝大多数分布于海拔1 000~1 800 m的山区, 陕西旬阳、湖南石门、河南商城、湖北英山等种群分布下限可低于1 000 m, 川西亚高山地区的宝兴种群分布上限超过1 800 m, 四川大学植物标本馆(SZ)00017835号标本记录显示可达2 700 m。生境为山坡林下阴湿处、沟谷溪边及悬崖峭壁60°左右的岩缝中, 生长状态多为散生。不少种群出现在耕地附近, 如四川万源、陕西略阳、甘肃武都、重庆南川和巫山种群, 生境破碎化严重。

|
图 1 巴山榧树地理分布 Fig.1 Geographical distribution of Torreya fargesii |
|
|

整合以植被覆盖图层为底层绘制的巴山地理分布图(图 2)与野外调查资料,结果表明,巴山榧树主要分布在常绿阔叶林、针阔叶混交林和常绿落叶阔叶混交林,少数分布点出现在灌丛、耕地及其他自然植被的镶嵌区中。实地调查结果显示,巴山榧树常以伴生种零星分布于群落中,林中常见裸子植物还有红豆杉(Taxus wallichiana var. chinensis)、南方红豆杉(T. wallichiana var. mairei)、三尖杉(Cephalotaxus fortunei)、粗榧(C. sinensis)、杉木(Cunninghamia lanceolata)和马尾松(Pinusmassoniana)等。以大娄山区的贵州柏枝山为例,巴山榧树分布在常绿落叶阔叶混交林,乔木层优势种有猴樟(Cinnamomum bodinieri)、香叶树(Linderacommunis)、黑壳楠(Lindera megaphyllas)、宜昌润楠(Machilus ichangensis)、山桃(Amygdalus davidiana)、山合欢(Albizzia kalkora)、八角枫(Alangium chinense)、盐肤木(Rhus chinensis)、漆树(Toxicodendron vernicifluum)和金佛山方竹(Chimonobambusa utilis)等,灌木层优势种有矮生栒子(Cotoneaster dammerii)、平枝栒子(C. horizontalis)、川莓(Rubussetchuenensis)、木莓(R. swinhoei)、长柄绣球(Hydrangea longipes)、圆锥绣球(H. paniculata)、毛叶楤木(Aralia chinensis var. dasyphylloides)、异叶梁王茶(Nothopanax davidii)、淡红荚蒾(Viburnumerubescens)、直角荚蒾(V. foetidum var. rectangutatum)、枇杷叶荚蒾(V. rhytidophyllum)和水马桑(Weigelajaponica var. sinica)等,草本层以菊科(Compositae)、百合科(Liliaceae)、毛茛科(Ranunculaceae)、禾本科和荨麻科(Urticaceae)植物为主。而重庆南川金佛山的巴山榧树则主要分布在灌丛,群落中悬钩子属(Rubus)、火棘属(Pyracantha)、栒子属(Cotoneaster)、花椒属(Zanthoxylum)、荚蒾属(Viburnum)、小檗属(Berberis)、忍冬属(Lonicera)和菝葜属(Smilax)植物占优势。

|
图 2 巴山榧树分布地植被覆盖 Fig.2 Landcover type in the distribution of Torreya fargesii |
14个环境因子共同进行模型运算所获得的基础溢出误差为5.45。选取最优模型时,依次去除14个环境因子所获得的溢出误差的95%置信区间的平均值列于表 2中,其中雨日频率、水汽压、年均温和汇流累积量被去除时,溢出误差明显降低(<5.45×0.95),而去除海拔、霜日频率、极端高温和极端低温则会使溢出误差明显升高(>5.45×1.05)。因此,雨日频率、水汽压、年均温和汇流累积量在最终的分析中被剔除,使用剩余10个环境因子建立1 000个模型并选择100个最优模型。以独立外部检验数据集为基础的卡方检验显示这100个模型在统计上都高度显著(P<0.01),即模型的预测能力远远好于随机模型(Kriticos et al.,2001)。最终获得的100个最优模型的溢出误差的平均值为4.29,远小于基础溢出误差。模型预测图与中国行政区图以及实际分布点叠加的结果见图 3,红、橙、绿、白4种颜色依次表示潜在分布的可能性由高到低。可以看出,我国的中部地区为巴山榧树的良好生长区(OV ≥90%),包括四川省东北部和西部,云南省东北部,重庆市东部,甘肃省东南部,陕西省南部,河南省南部,河南、湖北和安徽3省交界处,江西和湖南2省交界处,江苏、安徽和浙江3省交界处,贵州和湖北的大部分地区,以及吉林省的部分地区,大部分地区位于中亚热带和北亚热带,少数地区可至温带; 可生长区(70% ≤ OV<90%)和过渡区(50% ≤OV<70%)的范围非常狭窄,仅分布在良好生长区的边缘; 而我国的绝大部分地区是巴山榧树的不适生长区(OV<50%)。
|
|

|
图 3 生态位模型预测巴山榧树在我国的潜在分布区 Fig.3 Potential range prediction for Torreya fargesii applied GARP ecological niche model |
在以往的研究中,Desktop Garp经常被用于入侵生物学的预测(李双成等,2008; 陈立立等,2008; 余岩等,2009)。在筛选用于入侵生物的最优环境因子集合时,通常会采用一系列的刀切法分析,通过多次重复计算,每次省略一个环境参数,然后分析某一环境参数与遗漏误差(omission error)之间的相关性,确定最优的环境参数,即如果一个环境参数的存在与遗漏误差之间呈正相关关系,表明该环境参数会降低模型预测的准确性,该环境参数会在最终的分析中即被去掉。然而,这种方法并不适用于保护生物学的研究,最终预测的结果虽然能够包括所有预测点,但是却会使预测范围扩大化,这在入侵生物学的研究中并不会带来不良后果,因为入侵种往往具有较强的抗逆性,其真正可入侵范围可能会超出依据现有信息所做出的“准确”预测范围。对于保护生物学而言,需要被保护的物种的分布范围是在缩小的,这就要求预测范围更加精确,以便实施重点地段的就地保护和适生区域的迁地保护。因此,本研究在对环境因子进行筛选时,并未使用遗漏误差,而是使用了溢出误差。在Desktop Garp模型运算的结果列表中,溢出误差代表的是预测结果超出实际分布区所占的百分比。本研究在筛选最优环境因子集合时,进行了14次重复运算,每次省略1个环境参数,然后分析某一环境参数与溢出误差之间的相关性。如果某个环境参数的缺失会使溢出误差降低(小于基础溢出误差的95%),则表明该环境参数会扩大预测范围,降低预测精度,不应包括在最终的分析中。在表 2中,雨日频率、水汽压、年均温和汇流累积量最终被剔除,这样获得结果的精确度远好于使用全部14个环境因子所获得结果的精确度。
3.2 生态因子对巴山榧树的影响结合图 1和3可以发现,巴山榧树的分布预测区与我国海拔800~2 400 m的区域非常相近。在表 2中,海拔对于溢出误差的影响最大,因此我们推测,海拔是影响巴山榧树分布的最主要因素。程晓建等(2007; 2009)对榧树属其他种的研究表明,极端温度不低于-16 ℃时,榧树均可正常生长发育; 如果极端温度达-20 ℃以下的时间过长,榧树主梢常常会被冻死。在表 2中,极端低温、极端高温和霜日频率均是限制巴山榧树分布的主要环境因子,与事实相符。如果全球气候持续变暖,很多生物的最适海拔将发生上移,这对于像巴山榧树这类对海拔依赖的物种是极为不利的。最适海拔的上移将导致分布区的进一步破碎化,使得原本数量就不多的巴山榧树种群更难进行基因交流,可能对该物种的生存造成严重影响。榧树属植物幼年喜荫,高温和强光照会使幼苗枯死,一些中等高度的乔木可以为其提供遮光的环境; 而当苗木长大后,需光量逐渐增大,光照不足则生长不良。结实以后的榧树属植物需要更充足的光照以保证花芽分化和种子发育(高兆蔚,1997; 王昌腾,2005; 程晓建等,2007; 2009),因此,巴山榧树可能无法在过于高大的乔木林中生存。在对巴山榧树进行保护时,应尤其注意保护其生境,才能保证其正常的生长发育。值得注意的是,几个与水相关的环境因子—汇流累积量、水汽压、年降雨量,对巴山榧树的影响不大。虽然目前并未有对巴山榧树抗逆性的具体研究,但是对榧树属其他物种有过相关的报道,例如香榧(Torreya grandis ‘Merrillii’)就是抗旱性很强的树种,进入成年的香榧即使遇到夏、秋季长期干旱也很少发现落叶、枯枝和落果现象(程晓建等,2009)。巴山榧树对干旱的耐受性也很强,即使在悬崖、岩缝等无法保水的生境中也可以正常生长,这一特性非常有利于巴山榧树的人工繁育,巴山榧树可以作为我国中亚热带和北亚热带山区森林更新和荒山造林树种。
3.3 濒危原因推断物种濒危的原因是多方面的,有自然历史的原因,还有物种本身的原因以及人类的影响。虽然巴山榧树分布点和种群数量较多,但各种群的分布区域狭窄,生态条件恶劣,种群密度小。巴山榧树的分布格局与其他古老孑遗植物十分相似,呈岛屿化斑块状(高兆蔚,1997; 谢宗强等,1999; Schwartz et al.,2000; 徐晓婷等, 2008)。第四纪冰川时期,强烈的气候变迁造成了低海拔地区种群的灭绝,只留下呈间断分布的高海拔种群; 冰后期以来的气候波动则使已经片断化的种群数量进一步减少(谢宗强等,1999; 向巧萍,2001)。历史气候变迁可能是巴山榧树大尺度减少的主要原因。巴山榧树开花结实率低、天然林下幼苗稀少、生殖障碍大和更新困难也是其濒危的重要因素。
乱砍滥伐、盲目开荒和过度放牧等导致巴山榧树原生境破碎化,不少原有分布地已难觅巴山榧树踪迹,有的仅残存于采伐不可及的地方。在本次调查中,大多数巴山榧树种群及植株分布在地势险要的陡坡和悬崖峭壁上,各种群均不同程度地烙上了人类活动的印记,在林中、林缘、耕地边尚能见到伐木后留下的树桩或萌生的成丛嫩枝,尤其是位于矿区和耕地附近的种群及生境破坏更为严重。人为干扰促进了巴山榧树的濒危,甚至可能导致部分种群灭绝。
3.4 潜在分布区及保护策略从图 3可以发现,贵州省全境几乎都是巴山榧树的良好生长区(OV≥90%),海拔和植被分布也有相当大的范围契合。然而,在这一地区记录到的野生巴山榧树非常稀少,显然受到了极其严重的破坏,应当重点加强保护。在浙江,虽然目前还没有巴山榧树分布的报道,但根据本研究的预测,那里极有可能有巴山榧树分布。
濒危物种的保护主要有就地保护和迁地保护2种方法。巴山榧树水平分布范围跨度较大,分布点较多,应该加强现有巴山榧树种群的就地保护,减少人为破坏。由于巴山榧树生境特殊,所需生态条件苛刻,容易形成小种群,就地保护应从单一的物种保护转移到物种栖息地及生态系统的保护。然而从巴山榧树的地理分布和种群现状来看,仍有部分分布点位于自然保护区、风景名胜区或森林公园之外。随着集体林权制度的改革,林地使用权、林木所有权和经营权落实到户,广布于山区的珍稀保护植物命运堪忧。巴山榧树分布地的群众往往不知道该树种的确切名称,也不知其为国家重点保护植物,在择伐和间伐木材时易将其一同采集,一些群众还误将巴山榧树当作红豆杉采收卖给不法收购商。因此,在资源丰富的山区亟需加强生物多样性保护宣传。同时,建议林业部门组织力量对巴山榧树的分布及资源量做进一步考察,建立自然保护区或巴山榧树自然保护点,实施就地保护。对于生境退化严重、规模较小的种群采取迁地保护措施,并通过人工繁育巴山榧树幼苗在其适生区扩大种群规模。
本研究对巴山榧树的物种保护、资源普查和迁地保护等具有一定的指导意义。在资源本底调查和潜在分布区预测的基础上,应加强巴山榧树生物生态学、保护遗传学和人工繁育研究,为制订科学有效的保护措施提供理论依据。
| [] | 程晓建, 黎章矩, 戴文圣, 等. 2009. 香榧的生态习性及其适生条件. 林业科技开发, 23(1): 39–42. |
| [] | 程晓建, 黎章矩, 喻卫武, 等. 2007. 榧树的资源分布与生态习性. 浙江林学院学报, 24(4): 383–388. |
| [] | 陈立立, 余岩, 何兴金. 2008. 喜旱莲子草在中国的入侵和扩散动态及其潜在分布区预测. 生物多样性, 16(6): 578–585. |
| [] | 陈振德, 侯连兵, 谢立, 等. 2000. 榧子药材性状与商品鉴定. 中药材, 23(1): 19–21. |
| [] | 陈振德, 郑汉臣, 傅秋华, 等. 1998. 国产榧属植物种子油含量及其脂肪酸测定. 中国中药杂志, 23(8): 456–457. |
| [] | 丁宝章, 王遂义, 高增义. 1981. 河南植物志:第1册. 郑州, 河南人民出版社.. |
| [] | 傅书遐, 郑重, 王文采, 等. 2001. 湖北植物志:第1卷. 武汉, 湖北科学技术出版社.. |
| [] | 高兆蔚. 1997. 我国特有树种长叶榧树的生物学特性与保护问题研究. 生物多样性, 5(3): 206–209. |
| [] | 管中天. 1983. 四川植物志:第2卷. 成都, 四川人民出版社.. |
| [] | 郭泉水, 王春玲, 郭志华, 等. 2005. 我国现存梭梭荒漠植被地理分布及其斑块特征. 林业科学, 41(5): 2–7. DOI:10.11707/j.1001-7488.20050501 |
| [] | 康宁, 汤仲埙. 1995. 榧属分类学研究. 植物研究, 15(3): 349–362. |
| [] | 刘克明, 李丙贵, 刘林翰, 等. 2000. 湖南植物志:第2卷. 长沙, 湖南科学技术出版社.. |
| [] | 李双成, 高江波. 2008. 基于GARP模型的紫茎泽兰空间分布预测. 生态学杂志, 27(9): 1531–1536. |
| [] | 罗士德, 宁冰梅, 阮德春, 等. 1994. 红豆杉及其近缘植物中紫杉醇与同系物的高效液相色谱分析. 植物资源与环境, 3(2): 31–33. |
| [] | 苏应娟. 1997. 红豆杉科、三尖杉科和罗汉松科植物叶片结构的比较观察. 武汉植物学研究, 15(4): 307–316. |
| [] | 王昌腾. 2005. 野生长叶榧生物学特性与保护研究. 林业实用技术(10): 6–7. |
| [] | 王瑞. 2006. 我国严重威胁性外来入侵植物入侵与扩散历史过程重建及其潜在分布区的预测. 中国科学院研究生院博士学位论文.. |
| [] | 汪松, 解焱. 2004. 中国物种红色名录:第1卷. 北京, 高等教育出版社.. |
| [] | 汪小全, 舒艳群. 2000. 红豆杉科及三尖杉科的分子系统发育—兼论竹柏属的系统位置. 植物分类学报, 38(3): 201–210. |
| [] | 向巧萍. 2001. 中国的几种珍稀濒危冷杉属植物及其地理分布成因的探讨. 广西植物, 21(2): 113–117. |
| [] | 席以珍. 1986. 中国红豆杉科花粉形态的研究. 植物分类学报, 24(4): 247–252. |
| [] | 谢宗强, 陈伟烈. 1999. 中国特有植物银杉的濒危原因及保护对策. 植物生态学报, 23(1): 1–7. |
| [] | 徐晓婷, 杨永, 王利松. 2008. 白豆杉的地理分布及潜在分布区估计. 植物生态学报, 32(5): 1134–1145. |
| [] | 易同培, 杨林, 隆廷伦. 2006. 榧属(红豆杉科)一新种—四川榧. 植物研究, 26(5): 513–515. |
| [] | 余岩, 陈立立, 何兴金. 2009. 基于GARP的加拿大一枝黄花在中国的分布区预测. 云南植物研究, 31(1): 57–62. |
| [] | 于永福. 1999. 中国野生植物保护工作的里程碑:国家重点保护野生植物名录(第一批)出台. 植物杂志(5): 3–11. |
| [] | 张清华, 郭泉水, 徐德应, 等. 2000. 气候变化对我国珍稀濒危树种—珙桐地理分布的影响研究. 林业科学, 36(2): 47–52. DOI:10.11707/j.1001-7488.20000208 |
| [] | 张文驹, 陈家宽. 2003. 物种分布区研究进展. 生物多样性, 11(5): 364–369. |
| [] | 郑万钧, 傅立国. 1978. 中国植物志:第7卷. 北京, 科学出版社.. |
| [] | Adjemian J C Z, Girvetz E H, Beckett L, et al. 2006. Analysis of genetic algorithm for rule-set production (GARP) modelingapproach for predicting distributions of fleas implicated as vectors of plague, Yersinia pestis, in California. Journal of Medical Entomology, 43(1): 93–103. |
| [] | Attorre F, Francesconi F, Taleb N, et al. 2007. Will dragonblood survive the next period of climate change? Current and future potential distribution of Dracaena cinnabari(Socotra, Yemen). Biological Conservation, 138(3/4): 430–439. |
| [] | Beaumont L J, Pitman A J, Poulsen M, et al. 2007. Where will species go? Incorporating new advances in climate modelling into projections of species distributions. Global Change Biology, 13(7): 1368–1385. DOI:10.1111/gcb.2007.13.issue-7 |
| [] | Bell G. 2001. Neutral macroecology. Science, 293(5539): 2413–2418. DOI:10.1126/science.293.5539.2413 |
| [] | Fu L G, Li N, Mill R R. 1999. Taxaceae ∥Wu Z Y, Raven P H. Floraof China: 4.Beijing: Science Press. |
| [] | Funk V A, Zermoglio M F, Nasir N. 1999. Testing the use of specimen collection data and GIS in biodiversity exploration and conservation decision making in Guyana. Biodiversity and Conservation, 8(6): 727–751. DOI:10.1023/A:1008877222842 |
| [] | Institute of Gazetteer Research. 1994.Gazetteer of China. Beijing:China Map Press. |
| [] | Kriticos D J, Randall R P. 2001. A comparison of systems to analyse potential weed distributions∥Groves R H, Panetta F D, Virtue J G.Weed risk assessment. Victoria: CSRIO Publishing, 8(6): 61–79. |
| [] | Li J H, Davis C C, Donoghue M J, et al. 2001. Phylogenetic relationships of Torreya(Taxaceae)inferred from sequences of nuclear ribosomal DNA ITS region. Harvard Papers in Botany, 6(1): 275–281. |
| [] | Maurer B A. 2002. Biogeography: big thinking. Nature, 415(6871): 489–491. DOI:10.1038/415489a |
| [] | Nee S. 2002. Biodiversity: thinking big in ecology. Nature, 417(6886): 229–230. DOI:10.1038/417229a |
| [] | Schwartz M W, Hermann S M, van Mantgem P J. 2000. Estimating the magnitude of decline of the Florida torreya(Torreya taxifolia Arn). Biological Conservation, 95(1): 77–84. DOI:10.1016/S0006-3207(00)00008-2 |
| [] | Stockwell D, Peters D. 1999. The GARP modelling system: problems and solutions to automated spatial prediction. International Journal of Geographic Information Systems, 13(2): 143–158. DOI:10.1080/136588199241391 |
| [] | Zhang Y B, Ma K P. 2008. Geographic distribution patterns and status assessment of threatened plants in China. Biodiversity and Conservation, 17(7): 1783–1798. DOI:10.1007/s10531-008-9384-6 |