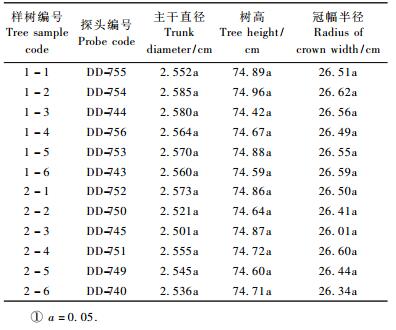
 2012, Vol. 48
2012, Vol. 48文章信息
- 李晓彬, 汪星, 汪有科, 张平
- Li Xiaobin, Wang Xing, Wang Youke, Zhang Ping
- 梨枣茎直径微变化的气象因子
- The Factors Influencing Stem Diameter Micro-Variation in Chinese Pear Jujube (Ziziphus jujuba)
- 林业科学, 2012, 48(1): 173-180.
- Scientia Silvae Sinicae, 2012, 48(1): 173-180.
-
文章历史
- 收稿日期:2011-01-17
- 修回日期:2011-11-23
-
作者相关文章



2. 西北农林科技大学 杨凌 712100;
3. 中国科学院研究生院 北京 100049
2. Northwest Agriculture and Forestry University Yangling 712100;
3. Graduate School of Chinese Academy of Sciences Beijing 100049
枣树(ziziphus jujuba)作为陕北黄土高原地区的主要经济林树种,因其可观的经济效益与生态效益,成为国家退耕还林(草)工程的优选树种与工程后续良性发展的优势树种。实施精确灌溉是现代节水农业的发展趋势,而掌握作物的需水信息是实现精确灌溉的基础和保障。为充分利用有限的水资源,建立节水高产的枣园灌溉制度,改变以往枣园传统的任意灌(水量)与随时灌(时间),必须确定枣树适宜的土壤水分与灌溉时间,即实现枣树的精确灌溉。植物茎直径微变化作为植物水分亏缺敏感性指标具有简单易行、对植株不具破坏性、可连续监测、自动记录并准确获取植物体内水分情况的优点(Turner,1987; Kozlowski et al., 1964; Namken et al., 1969; Huck et al., 1977); 但茎直径观测(trunk diameter measurements,TDM)容易受外界主要是气象因素影响,因此分析茎直径微变化的影响因子,探求准确、标准化监测信号的方法和指标,对于利用TDM指导灌溉具有重要意义(Goldhamer et al., 2004; Conejiero et al., 2007; García-Orellana et al., 2007; Velez et al., 2007)。国内对TDM指导灌溉的研究很少,尤其是对有关耐旱型品种枣树的研究未见报道。
已有研究表明:关键灌溉时期主要为发芽期、开花坐果期和果实膨大期,而果实膨大期灌水与否对最后的产量与果实品质有重要影响(康绍忠等,1996; 王中英,2000),如果果树在果实膨大期受到水分胁迫,将直接影响到产量与品质指标。因此本文重点研究临椅梨枣果实膨大期茎杆直径微变化的影响因子,通过设置2个土壤水势梯度,研究不同水势处理下枣树茎直径的动态变化规律,分析土壤水分与气象因子对茎直径微变化的影响,通过建立MDS对气象要素的回归模型,分析影响模型预测准确性的因素。为TDM在精确灌溉中的非正常波动提供不稳定因素分析,并为枣树基于植物自身水分状况需水信息的获取提供理论的精准性依据,为在枣树关键需水时期通过植株自身水分信息获取灌溉信息提供理论依据。
1 材料与方法 1.1 试验地概况试验在米脂县银州镇孟岔村山地微灌枣树示范基地防雨篷内进行。本区位于典型的黄土高原丘陵沟壑区(37.78° N,110.23° E,海拔870 m),属中温带半干旱性气候,气候干燥,昼夜温差大,日照丰富,适宜果树生长。年平均降雨量451.6 mm,主要集中在8—9月。试验地土质为黄绵土,密度1.38 g·cm-3,0 ~ 60 cm计划湿润层的田间持水量为26.35% (重量含水量),土壤表层水分稳定入渗速率为1.4 mm·min-1。
1.2 试验设计试验于2009年7月17日(第198天,2009年1月1日为第1天)开始,2009年8月16日(第228天)结束。供试材料为3年生临椅梨枣,采取小区试验,所选梨枣树形修剪均一,长势良好,平均主干茎直径2.39 cm (距地面20 cm处的茎秆数据),平均树高68.44 cm,平均冠幅半径25.94 cm。试验采用矮化树种,基本信息如表 1所示(枣树种是)。经统计分析试验样树的主干直径树高和冠幅半径无差异已有研究表明:土壤含水量在60%田间持水量以上时,适合果树生长(康绍忠等,1996; 王中英,2000),结合梨枣对于水分需求的特性和1 m土层内的田间持水量,本文将充分灌溉的土壤含水量始终控制在65%以上,当土壤含水量低于60%田间持水量时即视为水分亏缺。试验设置2个水分处理(充分灌溉和轻度亏缺)。T1:土壤水势控制在-15 ~-25 kPa(相对田持65% ~ 70%)范围内; T2:土壤水势控制在-45 ~-55 kPa(相对田持50% ~ 55%)范围内。每个处理自成一个小区,每个小区2行6棵树(每行3棵树),每棵树视为1次重复。采用水泥砌墙与周围土壤相隔并用塑料进行防渗,使小区土壤环境为封闭系统。各小区土壤为填充土,取自同一地点并经过均匀混合后按相同容重填到小区土槽内。
|
|
灌溉方式采用滴灌,为了保证灌溉的均匀性,一个小区2条毛管,每个毛管上等间距安装6个滴头,滴头流量为4 L·h-1。采用GP1自动控制灌溉,当小区土壤水势低于设定下限时,电磁阀将打开进行自动灌水,当土壤水势仪监测到的小区土壤水势达到设定上限时,即停止灌溉。若土壤水势仍低于设定的土壤水势下限,将继续灌溉直到满足设定的土壤水势范围。
1.3 观测指标与方法1) 土壤水势 用平衡式土壤水分张力计(EQ15: Equilibrium Tensiometer)监测土壤水势,测量范围为-1 500 ~ 0 kPa。每小区安装3个土壤水势仪探头,埋设深度均为30 cm,中间的水势仪探头与GP1相连,用来控制灌溉。其他探头与DL2e型数据采集器相连,每30 min自动记录一次数据。
2) 茎杆直径微变化 茎杆直径微变化采用DD型线性差分径向变化仪(简称LVDT,德国Ecomatik公司生产)连续测定。在每棵树安装1个探头,探头通过不胀钢框架安装在每株样树主干距地面15 cm处的北向,安装前先用木锉轻刮树干的死皮,以确保LVDT框架牢固和探头与主干接触良好,用隔热银箔纸将探头包住,以防止风、气温和降雨等对探头的直接影响。所有探头与DL2e型数据采集器(英国剑桥,Delta Device)相连,每30 min自动记录一次数据,直接采集的数据为MXTD(茎直径日最大值)与MNTD(茎直径日最小值),计算获取MDS(茎直径日最大收缩量,MXTD-MNTD)与DG(茎直径日生长量,MXTD-MXTD)。
3) 气象数据 距试验地10 m远的自动气象站按照国家气象局标准连续采集降雨量(mm)、太阳辐射(W·m-2)、气温(℃)、空气相对湿度(%)、2 m高处风速(m·s-1)和风向等。
① 参考作物蒸发蒸散量(ET0)和水汽压亏缺(VPD) 利用Penman-Monteith公式计算:
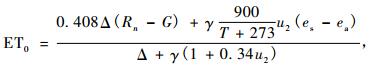
|
(1) |
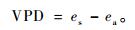
|
(2) |
式中: ET0为参考作物蒸发蒸散量,mm·d-1; VPD为饱和水汽压亏缺值,kPa; Rn为作物表面净辐射,MJ·(m-1 d-1); G为土壤热通量密度,MJ·(m-1 d-1); T为距地面2 m高处日平均气温,℃; u2为距地面2 m高处风速,m·s-1; es为饱和水汽压,kPa; ea为实际水汽压,kPa; Δ为水汽压曲线斜率,kPa ·℃; γ为湿热常数,kPa·℃。
② 大气水势Ψa 采用热力学化学势概念来计算:

|
(3) |
式中: Ψa为大气水势,kPa; T为日平均气温,℃; RH为日相对湿度,%。
2 结果与分析 2.1 土壤水势变化如图 1所示:试验期间T1处理的土壤水势始终维持在-15 ~-25 kPa,T2处理的土壤水势维持在-45 ~-55 kPa,并且T2与T1的土壤水势差值(Gap)稳定在-20 kPa左右,在0.05方差检验水平上,2个小区土壤水势的处理存在显著性差异。

|
图 1 土壤水势动态变化 Figure 1 Dynamic trend of soil |
试验期间,最高温出现在第204天为36.6 ℃,最低温出现在第223,224天,为13.3 ℃。平均温度为24.8 ℃,日温差最大值出现在第224天为22.5 ℃,最小值出现在第216天为3.5 ℃,平均日温差为13.1 ℃。
如图 2所示:试验期间,ET0,VPD与DTD (日温差) 3者具有较一致的变化趋势,而Ψa (大气水势)具有与3者反向的变化趋势。ET0,VPD与DTD分别出现6个波峰,7个波谷; 而Ψa出现7个波峰,6个波谷。ET0的最大值、最小值分别落在第1波峰和第5波谷上。VPD的最大值、最小值分别落在第2波峰和第5波谷上。DTD的最大值、最小值分别落在第6波峰和第4波谷上。Ψa的最大值、最小值分别落在第5波峰和第6波谷上。

|
图 2 ET0,VPD,DTD与Ψa随时间的动态变化 Figure 2 Dynamic of ET0, VPD, DTD and Ψa during the measurement period |
枣树的茎杆既具有运输水分的能力也具有储存水分的能力,前者通过木质部来实现,后者依赖茎杆中的薄壁细胞来完成。蒸腾开始时提供给叶子的水来自茎中的薄壁细胞(蒋高明等,2004)。白天茎中水的抽出使树干的直径不断发生变化,一般是早上最大而傍晚最小。茎杆收缩大多发生在木质部外围的活组织中,其细胞具有弹性较强的细胞壁,因而当水分抽出时细胞体积就将减小(蒋高明等,2004)。到了夜间蒸腾停止后,若土壤水分充足,根系吸收的水分来补充茎损失的水分使茎杆膨胀,茎杆复原或伴有生长; 反之,茎杆不能复原。茎杆直径日最大值(MXTD)与最小值(MNTD)之差即为茎杆最大日收缩量(MDS)。
茎直径变化包括因自然生长而导致的增粗和因为树木自身水分状况变化而引起的收缩或膨胀,如图 3所示。日变化过程可分为3个阶段: a)收缩阶段(Contraction phase) :茎直径从日最大值向其最小值变化的时间,一般从上午8 : 00—16 : 00; b)膨胀阶段(Expansion phase) :茎直径从日最小值恢复到前一天最大值所需的时间,一般从下午4点到次日凌晨2点; c)生长阶段(Stem increment phase) :茎直径从膨胀期结束至下一个日最大值的时间,一般从凌晨2点至8点。

|
图 3 茎直径日变化 Figure 3 Dvnamic of stem diameter during |
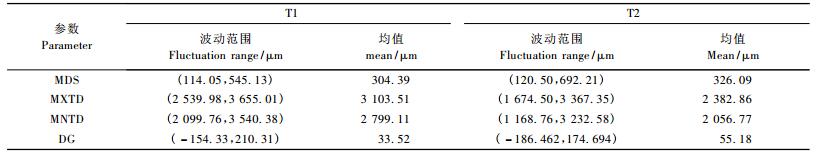
由表 2可知: T2处理的MDS值波动范围的上下限均大于T1处理的,均值比T1处理大21.7 μm; 而MXTD与MNTD值波动范围上下限均小于T1处理的,均值比T1处理的分别小720.65,742.34 μm。分析原因可能为:同一水平的大气外界条件下,水分供应充足,植物通过茎杆的根部提水量大,茎杆膨胀量大,表现为MXTD值大,随着大气蒸发强度增大,茎杆直径由膨胀转为收缩,在水分供应充足条件下,植物体地上部分(包括茎杆本身)会储存一部分水量,并且茎杆木质部始终有水分从根部输送到地上器官,使得茎杆收缩程度偏小,表现为MNTD值偏大; 而水分亏缺使得根部提水量小,表现为MXTD值小。当茎杆直径由膨胀转为收缩时,由于水分供应不足,所以茎杆收缩程度大,表现为MNTD值偏小。试验期间,T2处理的枣树DG(日茎直径增长量)均值要比T1处理的大21.66 μm,说明水分较充足条件下并不利于枣树的茎杆生长,而轻度的水分亏缺可以促进枣树的茎杆生长,这与枣树耐旱特性或许有关,试验结果与前人对枣树的茎秆微变化研究相符(张寄阳等,2006; 张平等,2010)。由于水分亏缺(T2)处理使得枣树茎杆直径的日生长量大于水分充足(T1)处理的,在一定程度上弥补了T2处理的MXTD值,综合T2处理的DG值大,水分亏缺造成的MNTD值偏小以及枣树根部提水后通过茎杆输送到地上器官水量的不稳定性,最终表现为T2的MDS(MXTD-MNTD)值波动范围与均值大于T1处理的。
|
|
如图 4所示: T1与T2处理具有相似的茎直径动态变化曲线,说明本试验水分处理之间,茎直径微变化变异性小,水分处理只是造成茎直径变化幅度的差异,而未改变茎直径的变化趋势,由于茎直径的变化主要是由气象因子与土壤水分综合作用引起的,由此得出枣树茎直径与气象因子之间具有很好的相关性。

|
图 4 MDS,MXTD,MNTD与DG随时间动态变化 Figure 4 Dynamic of MDS, MXTD, MNTD and DG during the measurement period |
试验期间,T2处理的MDS,DG值基本高于T1处理的,MDS之间的差异性显著时段为10天,DG之间的差异性显著时段为12天,但不存在明显规律。T1,T2的MDS最大值、最小值均在第204,216天,且2个处理的极值之间差异性显著。T1,T2的DG最小值均在第205天,最大值分别在第207,213天。结合气象因子分析: VPD与日均温最大值,ET0,DTD次最大值与Ψa次最小值出现在第204天。DTD,VPD与ET0最小值,Ψa最大值出现在第216天。由此说明: 1)不同水分处理下,MDS在遇到极端气象条件下会发生巨大波动或出现极值,而在极端气象条件下,水分处理造成MDS极值间出现显著性差异。综合分析: MDS可以作为水分敏感性指标指示枣树自身水分状况。2)极端气象条件会减弱枣树茎杆直径的生长量,而不同水分处理下的减弱程度不存在显著性差异。综合分析:适宜的水分亏缺可以促进枣树茎直径增长,但不显著。
试验期间,T1与T2处理的MXTD,MNTD随时间都具有增大的趋势,并且处理下的动态变化趋势相类似。MNTD呈现波折型动态变化,MXTD则呈现平缓型动态变化,MNTD的波动强度大于MXTD。分析原因可能为:一是MXTD主要出现在早晨6点前后,此时的每日气象因素差异不显著,变动幅度小; 而MNTD主要出现在下午4点前后,此时每日的气象因素差异较显著,变动幅度大。二是枣树茎杆直径的日生长阶段与MXTD同时段,DG在一定程度上掩盖了外界对MXTD的影响,显现出MXTD呈平稳型动态变化。T1,T2处理下MXTD,MNTD差异性显著时段分别为第18,20天。在第31天的试验处理期间,MXTD,MNTD较好的表现出了枣树自身水分状况,可以作为枣树水分敏感性指标指示枣树自身水分状况与土壤水分情况。在第220天以后,T1与T2处理的MXTD,MNTD之间差异性由显著变为不显著,差异呈现逐渐减小趋势。根据以往研究分析原因可能为:一是枣树在第220天(8月8号)以后,叶面积与新梢不再增长(碳代谢发生变化),生命活动减弱,蒸腾量减少,对水分的需求减小。二是第220天以后,气象因素在这个时段内出现极大的突变,为消减外界对枣树作用的激变,枣树减弱了自己活动量。
2.4 茎直径日最大收缩量(MDS)与气象因子的回归分析植物茎中储存的水分可以用于蒸腾过程中临时用水之需(蒋高明等,2004),所以植物茎杆直径的微变化可以传递出自身的水分信号。茎杆最大日收缩量(MDS)是茎杆直径微变化1个重要的指标,受土壤水分和气象要素的综合影响(张寄阳等,2006; 孟兆江等,2006)。
如图 5所示:不同处理下MDS与气象因子之间具有很好的一元二次曲线拟合性(判定系数r2在0.001水平上差异显著)。T1处理的MDS与各气象因子的拟合度均要高于T2处理的,由此说明,在同等大气外界条件下,水分供应充足的枣树MDS与气象因素具有较高的相关性,而水分亏缺则会影响MDS受气象因素综合作用的影响。T1处理的MDS与ET0的拟合度最高,与DTD,VPD的拟合度次之,与Ψa的拟合度最低。T2处理的MDS与DTD拟合度最高,与VPD,ET0的拟合度次之,与Ψa的拟合度最低。与VPD,ET0较高的拟合度说明了MDS在一定程度上表现了枣树蒸腾流的强度。DTD与2种水势处理下的MDS都具有较高的拟合度,分析原因可能为:温度通过对根系活力、茎杆木质部组织活性以及水分能态产生影响进而影响到MDS的变化,虽然温度不是一个精确的大气蒸发强弱的指标,但由于温度作为气象因子可方便容易的被观测,所以温度是MDS一个很好的预测指标。

|
图 5 MDS与气象因子间的相关分析 Figure 5 Relationships between MDS and meteorological factors |
MDS作为枣树水分亏缺敏感性指标,受土壤水势和气象因子综合作用的影响,可以反映土壤水分状况与枣树自身水分情况,通过建立MDS对气象因子的多元线性回归模型来预测枣树MDS动态变化,进而反映枣树生长健康状况与水分亏缺程度。通过分析影响模型预测准确性的因素对模型加以修正。
如表 3所示: TI,T2处理的MDS与气象因子之间都具有很好的线性拟合度。并且T1处理的线性拟合程度要大于T2处理的。如图 6所示: T1处理的MDS实测值偏离预测值较大的出现在第203,204,205,213,214天,T2处理的MDS实测值偏离预测值较大的出现在第198,203,204,213,214天。结合气象因子分析,VPD与日均温最大值,ET0,DTD次最大值与Ψa次最小值出现在第204天,说明试验期间的极端气象因素会影响到MDS预测的准确性,且T1,T2的DG最小值均在第205天,最大值分别在第207,213天。由此也说明枣树自身生长的极值点也会影响到MDS预测的准确性。
|
|
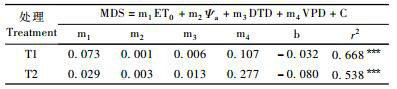

|
图 6 MDS预测值与实测值的分布 Figure 6 Distributing of measured value and predictive value to MDS |
1) 土壤水分基础值不同,枣树生命活动强度以及对土壤水分影响程度也不同。由图 1可以看出:在水分供应充足条件下,枣树生命进程对土壤水分变化的影响小,土壤水势呈平稳型动态变化; 而在水分相对不足条件下,枣树生命进程对土壤水分变化的影响大,土壤水势呈波动型动态变化。这与SPAC理论中,土壤水作为植物水分的主要源泉,大气植物蒸腾耗水量可以间接影响到土壤含水量的变化是相吻合的。反观土壤水势的不同,枣树茎直径的微变化(MDS,MXTD,MNTD,DG)出现差异性。说明土壤—植物界面的水分动态变化对植物生命活动产生影响。这与Goldhamer等(2004)指出在某种土壤湿润情况下的水分保证植物对水分的所需程度,只有植物本身才能给出正确和精确的回答,也就是植物在土壤水分动态变化过程中能够作出响应,植物的形态与生理状况可以作为衡量土壤水分适宜程度标志的观点具有一致性。从植物自身角度获取土壤水分情况与植物自身水分状况信息的方法是可行的。
2) 茎直径微变化受土壤水分与气象因子的综合影响,茎直径各参数受影响程度不同,具有一定差异性。水分不足在一定程度上增大了枣树茎直径最大日收缩量(MDS)与日生长量(DG),减小了枣树茎直径日最大值(MXTD)与日最小值(MNTD)。土壤水分不同,极端气象因素(试验期间的气象极值)会造成MDS出现极值并且差异显著,而对DG影响不显著。说明MDS可以作为枣树水分敏感性指标指示土壤水分情况与植物自身水分状况。而在试验期间,MXTD与MNTD由差异显著到不显著可能说明了随着枣树生命活动量(叶面积与新梢不在变化)的减弱,体内碳代谢发生变化,枣树自身对水分需求量减小,进而一定程度上消减了大气蒸发对茎直径的影响。由试验前期T2处理的MDS大于T1处理的到后期小于T1处理的MDS,由此说明,MDS受植物生育期内生命活动量的影响,MDS也可以作为植物碳状况变化的一个内容。这与Goldhamer等(1999)对桃树茎直径变化规律研究结果相符。根据枣树生育期分阶段茎直径受土壤水分与气象因子影响的程度与状况有待进一步研究探讨。
3) 茎直径日最大收缩量(MDS)与VPD,ET0密切相关,说明MDS受大气蒸发强度影响较大,表征了蒸腾流的强弱。与DTD密切相关,分析原因可能为,温度通过对根系活力、茎杆木质部组织活性以及水分能态产生影响进而影响到MDS的变化,虽然温度不是一个精确的大气蒸发强弱的指标,但由于温度作为气象因子可方便容易的被观测,所以温度是MDS一个很好的预测指标。通过建立MDS对气象因子的回归模型,发现MDS对气象因子有较好的多元一次线性回归性。但极端气象条件会影响到预测的准确性。这可能与枣树的生命机制有关,具体原因以及极端气象的范围有待进一步研究。国外研究方面,Ortuño等(2009)和Moriana等(2010)研究认为: MDS变化与树龄有关Marsal等(2002)、Intrigliolo等(2001)、Moriana等(2010)认为MDS变化还与物侯期和生育期有关,果实的生长、成熟和采摘直接影响到MDS的变化。所以分生育期不同阶段MDS对不同气象因子的相关性大小以及拟合形式有待进一步探讨。
本试验由于水分处理较少,气象条件变化大,试验条件不易控制等局限性,结论的可靠性或许会受到影响,但通过严格控制边界条件,消减试验误差,试验结论具有一定参考价值,对枣树茎直径微变化的水分、气象影响因素,尚需要通过多水分梯度处理进行深入研究。
蒋高明, 常杰, 高玉葆, 等. 2004. 植物生理生态学[M]. 北京: 高等教育出版社.
|
康绍忠, 蔡焕杰. 1996. 农业水管理学[M]. 北京: 中国农业出版社.
|
孟兆江, 段爱旺, 刘祖贵, 等. 2006. 温室茄子茎直径微变化与作物水分状况的关系[J]. 生态学报, 26(8): 2516-2522. |
王中英. 2000. 果树抗旱生理[M]. 北京: 中国农业出版社.
|
张平, 汪有科, 湛景武, 等. 2010. 红枣开花坐果期水分信息诊断指标适宜性分析[J]. 灌溉排水学报, 29(3): 94-97. |
张寄阳, 段爱旺, 孟兆江, 等. 2006. 基于茎直径微变化的棉花适宜灌溉指标初步研究[J]. 农业工程学报, 22(12): 86-89. DOI:10.3321/j.issn:1002-6819.2006.12.017 |
Conejero W, Alarcón J J, Garcia-Orellana Y. 2007. Evaluation of sap flow and trunk diameter sensors for irrigation scheduling in early maturing[J]. Tree Physiol, 27(12): 1753-1759. DOI:10.1093/treephys/27.12.1753 |
García-Orellana Y, Ruiz-Sánchez M C, Alarcón J J, et al. 2007. Preliminary assessment of the feasibility of using maximum daily trunk shrinkage for irrigation scheduling in lemon trees[J]. Agric Water Manag, 89(1 /2): 167-171. |
Goldhamer D A, Fereres E, Mata M, et al. 1999. Sensitivity of continuous and discrete plant and soil water stress status monitoring in peach trees subjected to deficit irrigation[J]. J Am Soc Hortic Sci, 124(4): 437-444. |
Goldhamer D A, Fereres E. 2004. Irrigation scheduling of almond trees with trunk diameter sensors[J]. Irrig Sci, 23(1): 11-19. DOI:10.1007/s00271-003-0088-0 |
Huck M G, Kleper B. 1977. Water relations of cotton[J]. Ⅱ. Continuous estimates of plant water potential from stem diameter measurements. Agronomy Journal, 69(4): 593-597. |
Intrigliolo D S, Castel J R. 2007. Crop load affects maximum daily trunk shrinkage of plum trees[J]. Tree Physiology, 27(1): 89-96. DOI:10.1093/treephys/27.1.89 |
Kozlowski T T, Winget C H. 1964. Diurnal and seasonal variation in radii of tree stems[J]. Ecology, 45(1): 149-155. DOI:10.2307/1937115 |
Marsal J, Gelly M, Mata M, et al. 2002. Phenology and drought affects the relationship between daily trunk shrinkage and midday stem water potential of peach trees[J]. Journal of Horticultural Science & Biotechnology, 77(4): 411-417. |
Moreno F, Conejero W, Martin-Palomo M J, et al. 2006. Maximum daily trunk shrinkage reference values for irrigation scheduling in olive trees[J]. Agricultural Water Management, 84(3): 290-294. DOI:10.1016/j.agwat.2006.03.005 |
Moriana A, Giron I F, Martin-Palomo M J, et al. 2010. New approach for olive trees irrigation scheduling using trunk diameter sensors[J]. Agricultural Water Management, 97(11): 1822-1828. DOI:10.1016/j.agwat.2010.06.022 |
Namken L N, Bartholic J F, Runkles J R. 1969. Monitoring cotton plant stem radius as an indication of water stress[J]. Agronomy Journal, 61: 891-893. DOI:10.2134/agronj1969.00021962006100060018x |
Ortuño M F, Brito J J, García-Orellana Y, et al. 2009. Maximum daily trunk shrinkage and stem water potential reference equations for irrigation scheduling of lemon trees[J]. Irrig Sci, 27(2): 121-127. DOI:10.1007/s00271-008-0126-z |
Turner N C. 1987. Crop water deficits: A decade of progress[J]. Adv in Agron, 39: 1-51. |
Velez J E, Intrigliolo D S, Castel J R. 2007. Scheduling deficit irrigation of citrus trees with maximum daily trunk shrinkage[J]. Agric Water Manag, 90(3): 197-204. DOI:10.1016/j.agwat.2007.03.007 |