 2012, Vol. 48
2012, Vol. 48文章信息
- 刘亚梅, 刘盛全
- Liu Yamei, Liu Shengquan
- 人工倾斜火炬松3年生苗木应压木的解剖性质
- Anatomical Properties of Compression Wood of Three-Year-old Loblolly Pine Induced by Artificial Inclination
- 林业科学, 2012, 48(1): 131-137.
- Scientia Silvae Sinicae, 2012, 48(1): 131-137.
-
文章历史
- 收稿日期:2010-04-09
- 修回日期:2010-04-28
-
作者相关文章



应力木是树木中的非正常木材组织,通常是指在外力作用下而形成的弯曲树干或树枝试图恢复到它原来位置,从而形成了解剖和物理力学性质明显不同的木材(IAWA Committee,1989)。在裸子植物中,应力木通常出现在倾斜树干或树枝的下部,被称为应压木; 在被子植物中,应力木通常出现在倾斜树干或树枝的上部,称为应拉木。由于应力木与正常木解剖、化学、物理、力学性质以及加工利用特性明显不同,所以应力木被国际木材学界公认为是影响木材品质和加工利用的主要问题之一(Panshin et al., 1980; Zobel,1989; Thibaut et al., 2001)。
木材解剖性质对木材物理、力学和化学性质有很大的影响,是研究木材材性的基础。应压木一般比正常木管胞的次生壁厚、管胞长度短(Onaka,1949; Wardrop et al., 1950)、次生壁的微纤丝角度大、木素含量高(Wilson et al., 1977; Timell,1986),且应压木具有胞间隙和螺纹裂隙(Côte et al., 1965; Yumoto et al., 1983)。尽管国内外对应压木解剖性质有不同程度的认识和研究,但大多集中在对成熟林分中倾斜树干应压木性能的测试方面,而对采用人工倾斜苗木树干来研究应压木的木材性质变化规律在国内还鲜见报道。
本文以盆栽的火炬松(Pinus taeda) 3年生苗木为研究对象,人工控制其倾斜角度(不是局部授力),测定不同倾斜角度下火炬松苗木木质部应压木的解剖性质,分析其变化规律,为应压木形成理论的探讨和其木材的合理利用提供科学依据。
1 材料与方法 1.1 试验材料2007年4月初,在安徽农业大学林学与园林学院内的实验苗圃地内种植了60株大小一致的3年生火炬松苗木(苗木高度为75 cm,地径为2.5 cm)。苗木分5组栽植在营养盆内,每组12株。2007年5月,将5组苗木倾斜,与垂直方向成不同的角度:0°,15°,30°,45°,60° (图 1),其中0°为对照组,15°~ 60°为应压木组。为了避免外界对树木生长的干扰,每株苗木自地面往上用宽的塑料带与一根2 m长的竹竿固定在一起,使树干保持5种倾斜角度。

|
图 1 苗木人工倾斜示意 Figure 1 Schematic diagram for artificial inclination of seedlings |
火炬松生长4个月后(地径为2.78 ~ 2.85 cm),2007年9月开始每隔3个月测定树干的生长应变,从中选取5株,在树干1 /3高度处截取轴向2 cm长的幼茎作为试样。试样先根据偏心程度分为应压木区和对应木区,应压木区位于倾斜树干的下部,年轮较宽; 对应木区位于倾斜树干的上部,年轮较窄。本文所选试样区域为应压木区,先在应压木区年轮最外层切取1 cm × 1 cm的试块,后将试块通过软化处理,再在回转式切片机上切成10μm厚横切面切片和20 μm厚弦切面切片,分别用于火炬松应压木各种不同解剖性质的测定分析。测量部位尽量在年轮最外围区域,保证是次生木质部的应压木区域。
1.2.2 测量方法应压木显微构造特征观测是将横切面切片,经染色后放在显微图像分析系统中观察。超微构造特征观测是将先将横切面切片加入乙醇逐级梯度脱水,脱水的乙醇浓度依次为20% →40% →60% →80% →95% →100% (2次),切片在每级停留30 min; 然后将少量导电胶涂到样品托上,用镊子轻夹切片侧面,保证横切面向上贴牢在涂胶上; 粘贴后,待导电胶干透,进行真空干燥约3 min,将样品托置于场发射扫描电镜(FE-SEM) S-4800(Hitachi)内进行喷金处理后在图像软件中观察。管胞长度的测定是将试样离析成单根纤维后,再在显微投影仪上测得。管胞宽度、双壁厚和组织比量测量是将横切面切片,经染色后放在显微图像分析系统中测量。微纤丝角度的测定方法是将弦切面切片,经脱木素、染色和固定处理后,使用偏光显微镜测量。
1.2.3 数据处理方法采用Excel,Photoshop及SPSS软件对数据进行分析处理。
2 结果与分析 2.1 不同倾斜角度下火炬松应压木管胞构造特征变化规律 2.1.1 管胞显微构造特征图 2为不同倾斜角度火炬松横切面晚材管胞的显微构造特征。从图 2可以看出: 0°时,管胞壁厚度较薄,平均厚度为2.20μm,管胞形状为多边形,胞间隙未见; 15°时:管胞开始具有应压木的特征,管胞壁厚度增加,平均厚度为2.32 μm,管胞形状由多边形向圆形转变,胞间隙可见; 30°时,管胞具有明显的应压木特征,管胞壁厚度较厚,平均厚度为2.51 μm,管胞形状大多为圆形,胞间隙较大; 45°时,管胞应压木特征非常明显,管胞壁厚度最大,平均厚度为2.70 μm,管胞形状近似圆形和椭圆形,胞间隙很大; 60°时,管胞应压木特征较明显,管胞壁厚度较厚,平均厚度为2.62 μm,管胞形状近似圆形和椭圆形,胞间隙较大。总体来看,火炬松直立生长树干(0°)管胞显微构造为典型正常木结构,随着倾斜角度的增加,应压木特征越来越明显,管胞壁逐渐变厚,管胞形状逐渐变圆,胞间隙明显增大,其中管胞壁厚度在倾斜45°时达到最大。本试验观测结果与Yamaguchi等(1983)对日本落叶松(Larix kaempferi)人工倾斜后所形成的应压木管胞特征以及Warensjö(2003)和Burgert等(2004)对几种不同树种应压木管胞特征的描述相类似。

|
图 2 不同倾斜角度应压木管胞显微构造特征 Figure 2 Microstructure images of compression wood tracheid at different angles |
在显微构造特征测定分析的基础上,本文又进一步观测了不同倾斜角度火炬松应压木横切面晚材管胞超微构造特征,特别是胞壁螺纹裂隙的变化情况。图 3为不同倾斜角度管胞超微构造特征,表 1为不同倾斜角度胞间隙和螺纹裂隙的特征值记载。从图 3和表 1可以看出: 0°时,管胞壁薄,形状为多边形,胞间隙未见,管胞壁螺纹裂隙不明显; 15°时,管胞壁较厚,形状稍圆,胞间隙可见,管胞壁具有较浅的螺纹裂隙; 30°时,管胞壁厚,形状较圆,胞间隙增多,管胞壁具有较深的螺纹裂隙; 45°时,管胞壁厚,形状较圆,胞间隙明显,管胞壁具有螺纹裂隙,且裂隙较宽、较深; 60°时,管胞壁厚,形状较圆或多角形,胞间隙较大,管胞壁螺纹裂隙明显,且裂隙较宽、较深。总体而言,直立生长的火炬松超微构造为正常木特征,随着倾斜角度的增加,火炬松苗木应压木的管胞壁逐渐变厚,管胞形状变圆,胞间隙增多、增大,螺纹裂隙变宽、加深,特别是倾斜30°以后,胞壁螺纹裂隙变宽、加深非常明显。Côte等(1965)和Yumoto等(1982)研究表明:应压木是由壁厚且圆的管胞和胞间隙组成,且管胞具有螺纹裂隙; Yoshizawa等(1987)和Timell(1983)也在裸子植物树种的应压木中观察到螺纹裂隙。本研究发现:不同倾斜角度下的应压木管胞中不仅具有螺纹裂隙,而且螺纹裂隙还随着倾斜角度的变化而变化。

|
图 3 不同倾斜角度应压木管胞超微构造特征 Figure 3 FE-SEM images of compression wood tracheid at different angles a.横切面管胞Tracheid of cross section; b.管胞壁内Inner tracheid cell wall. |
|
|
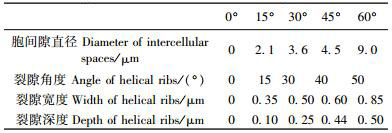
针叶材的管胞形态特征是判定其材性的重要指标,本文研究的管胞形态特征主要包括管胞长度、管胞宽度、双壁厚。
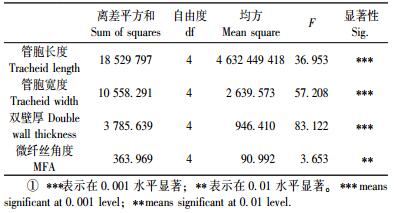
2.2.1 管胞长度不同倾斜角度火炬松应压木管胞长度的平均值和变异系数如表 2所示。从表 2可知:不同倾斜角度的火炬松管胞长度分布在1 189 ~1 506 μm之间,变异系数为6.2% ~ 10.6%,其中不同倾斜角度应压木管胞长度平均值比正常木短18%。Onaka(1949)和Wardrop等(1950)研究表明:应压木的管胞长度比正常木短10% ~ 40%;Ollinmaa(1981)也得出应压木管胞长度比正常木管胞长度短。本结果与之相一致,这是因为应压木区形成层原始细胞分裂加快会导致细胞长度的变短(Wardrop et al., 1950)。图 4a为管胞长度随倾斜角度变化情况,从图中可以看出:管胞长度随倾斜角度增加逐渐下降,其中30°时下降最大,说明30°为管胞长度变化的一个转折点,在30°时形成层原始细胞分裂最快。本结果与Yamashida等(2009)对不同倾斜角度的日本柏树(Chamaecyparis obtusa)管胞长度的描述相一致。不同倾斜角度管胞长度的方差分析(表 3)表明:倾斜角度对火炬松管胞长度的影响在0.001水平达极显著。
|
|

|
|

|
图 4 不同倾斜角度应压木胞形态特征和微纤丝角度的变化 Figure 4 Variation of trachied morphology and MFA of compression wood at different inclination angles a.管胞长度Tracheid length; b.管胞宽Tracheid width; c.双壁厚Double wall thickness; d.微纤丝角度MFA. |
由表 2可知:不同倾斜角度火炬松苗木的管胞宽度平均值为25.24 ~27.84 μm,变异系数为6.2% ~ 11.9%,其中不同倾斜角度应压木管胞宽度均值比正常木小6.3%。图 4b为不同倾斜角度管胞宽度的变化情况,从图中可以看出:管胞宽度随倾斜角度增加先减小后又略有增加,45°达最小值。不同倾斜角度管胞宽度的方差分析(表 3)表明:倾斜角度对火炬松管胞宽度的影响在0.001水平达极显著。
双壁厚平均值为4.40 ~ 5.41 μm (表 2),变异系数为8.4% ~ 18.9%,不同倾斜角度应压木双壁厚比正常木大11%。图 4c为双壁厚随倾斜角度的变化情况,从图中可以看出:双壁厚随倾斜角度增加先增大后减小,且在45°时达最大值。不同倾斜角度双壁厚的方差分析(表 3)表明:倾斜角度对火炬松双壁厚的影响在0.001水平达极显著。
2.3 不同倾斜角度下火炬松应压木微纤丝角度变化规律微纤丝角是次生壁S2层中微纤丝排列方向与细胞轴向之间的夹角,是细胞壁的基本性质之一,也是重要的木材解剖性质。从不同倾斜角度微纤丝角度变化(表 2)可以看出:火炬松应压木微纤丝角度都较大,其值分布在29.1° ~ 32.2°之间,变异系数为7.6% ~ 10.8%,不同倾斜角度应压木微纤丝角度均值比正常木大6%,说明了应压木S2层微纤丝角度比正常木大,此结果与Wilson等(1977)和Timell(1986)报道结果相类似。图 4d为不同倾斜角度微纤丝角度的变化情况,从图中可以看出:微纤丝角度随倾斜角度增加而增大后基本保持不变,30°达最大值,45°和60°值与15°值相近。不同倾斜角度微纤丝角度的方差分析(表 3)表明:倾斜角度对火炬松微纤丝角度的影响在0.01水平达显著。
2.4 不同倾斜角度下火炬松应压木组织比量变化规律木材组织比量是指各种木材构造分子所占的百分比,是反映各种木材构造分子定量变化的重要指标。Panshin等(1980)描述针叶材主要构造分子所占的比例为:轴向管胞90% ~ 95%,木射线6% ~ 9%,轴向树脂道0.1% ~ 1%。本文主要对不同倾斜角度下火炬松苗木的轴向管胞、木射线和轴向树脂道的组织比量进行了测定,结果如表 4所示,从表 4可知:不同倾斜角度管胞比量为90.9% ~ 91.8%,变异系数为1.1% ~ 1.7%,不同倾斜角度应压木管胞比量均值比正常木少3%;木射线比量变化幅度为7.0% ~7.7%,变异系数为5.8% ~ 10.1%,而不同倾斜角度射线比量均值与正常木相似,说明木射线组织比量变化不大,Panshin等(1980)也指出在同一树种内应压区木射线以及射线细胞与正常木基本上没有大的区别。树脂道比量分布范围为0.9% ~ 1.5%,变异系数为5.9% ~ 21.1%,不同倾斜角度应压木树脂道比量均值比正常木多31%,说明应压木在其形成过程中泌脂细胞明显增多。王婉华等(1991)对火炬松幼龄材进行研究,测定其树脂道比量为1.29%,本文树脂道比量大小与之相一致。
|
|
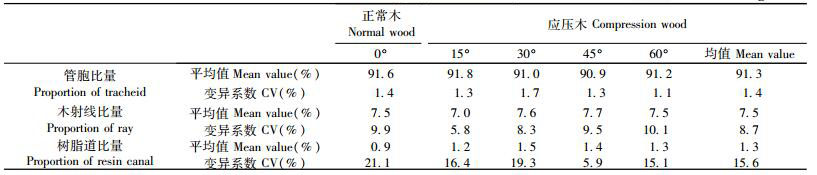
图 5为不同倾斜角度各组织比量的变化情况,从图中可以看出:随着倾斜角度的增加,管胞比量没有统一的变化模式,呈波动变化; 木射线比量同样是呈波动变化; 树脂道比量先增大后略有减小,30°达最大值,说明不同倾斜角度下应力木组织比量变化非常大,但组织比量的大小除受倾斜角度的变化影响外,还受其他因子的影响。

|
图 5 不同倾斜角度应压木组织比量的变化 Figure 5 Variation of tissue proportion of compression wood at different inclination angles a.管胞比量Proportion of tracheid; b.木射线比量Proportion of ray; c.树脂道比量proportion of resin canal. |
不同倾斜角度组织比量的方差分析如表 5所示,从表中可以看出:倾斜角度对火炬松树脂道比量的影响在0.001水平达极显著,而倾斜角度对火炬松管胞比量和木射线比量的影响均不显著。
|
|
本文以3年生火炬松苗木为研究对象,在生长季节开展人工倾斜树干试验,将苗木分成5组(每组12株),与垂直方向倾斜成不同的角度(0°,15°,30°,45°,60°),苗木生长4个月后开始测定不同倾斜角度下火炬松应压木的解剖性质,分析不同倾斜角度下火炬松应压木解剖性质的变化规律,得出以下结论: 1)随着倾斜角度的增加,管胞壁变厚,管胞形状变圆,胞间隙逐渐增多、增大,螺纹裂隙逐渐变宽、加深,其中30°以后应力木典型特征非常明显。2)随着倾斜角度的增加,管胞长度先减小后增大,30°达最小值; 管胞宽度先减小后增大,45°达最小值; 双壁厚先增大后减小,45°达最大值。方差分析表明不同倾斜角度对管胞长度、宽度及双壁厚影响均极显著。3)随着倾斜角度的增加,微纤丝角度先增大后减小,30°达最大值。方差分析表明不同倾斜角度对微纤丝角度影响显著。4)随着倾斜角度的增加,管胞比量及木射线比量呈波动变化; 树脂道比量先增大后减小,30°达最大值。方差分析表明不同倾斜角度对树脂道比量影响显著,而对管胞比量及木射线比量影响不显著。5)从不同倾斜角度下各种解剖性质的变化趋势来看,30° ~ 45°为应压木解剖性质形成的重要转折点,具体原因还需进一步研究。
综上所述,火炬松经过人工倾斜后,直立生长与倾斜生长的树干呈现不同的解剖特征,直立生长的树干(0°)具有正常木特征,倾斜生长的树干(15° ~60°)具有明显的应压木特征。不同倾斜角度对火炬松应压木管胞长度、管胞宽度、双壁厚、微纤丝角度、树脂道比量影响均显著; 且在本研究的5种不同倾斜角度中,30° ~ 45°是应压木各种解剖性质形成的转折点,但是从超微观特征可以看出,随倾斜角度的增加胞间隙逐渐增多、增大,螺纹裂隙逐渐增多、变宽、加深。造成这种现象的原因,需要在后续的研究中进一步探讨。本研究在此基础上连续跟踪测定了不同倾斜角度火炬松应压木的形成过程,后续还将开展不同倾斜时间下火炬松应压木每组角度解剖性质的变化情况,与本文结果进行整体对照分析后,将得到更加完善的结论。
王婉华, 阮锡根, 潘彪, 等. 1991. 湿地松, 火炬松和马尾松幼龄材构造特征的研究[J]. 南京林业大学学报, 15(4): 89-92. |
Burgert I, Frühmann K, Keckes J, et al. 2004. Structure-function relationships of four compression wood types: micromechanical properties at the tissue and fibre level[J]. Trees, 18(4): 480-485. |
Côte W A, Day A C. 1965. Anatomy and ultrastructure of reaction wood∥Côte W A. Cellular ultrastructure of woody plants. Syracuse University, N Y, 391-418.
|
IAWA Committee. 1989. IAWA list of microscopic features for hardwood identification[J]. IAWA Bull, 10: 219-332. DOI:10.1163/22941932-90000496 |
Ollinmaa P J. 1981. On certain physical properties of wood crown on drained swamps[J]. Silva Fennica, 15(3): 347-358. |
Onaka F. 1949. Studies on compression and tension wood. Wood Research Institute, Kyoto University, Kyoto, Japan.
|
Panshin A J, De Zeeuw C. 1980. Textbook of wood technology[M]. 4th ed. New York, N Y: McGraw-Hill.
|
Thibuat B, Gril J, Fournier M. 2001. Mechanics of wood and trees: some new highlights for an old story[J]. Comptes Rendus de I'Académie des Sciences-Series ⅡB-Mechanies, 329(9): 701-706. DOI:10.1016/S1620-7742(01)01380-0 |
Timell T E. 1983. Origin and evolution of compression wood[J]. Holzforschung, 37(1): 1-10. DOI:10.1515/hfsg.1983.37.1.1 |
Timell T E. 1986. Compression wood in Gymnosperms[M]. Berlin: Springer-Verlag.
|
Wardrop A B, Dadswell H E. 1950. The nature of reaction wood, Ⅱ: the cell wall organization wood tracheid[J]. Aust J Sci Res Ser B, Biol Sci, 3(1): 1-13. |
Warensjö M. 2003. Compression wood in Scots pine and Norway spruce———distribution in relation to external geometry and the impact on dimensional stability in sawn wood[J]. Doctoral Thesis, Swedish University of Agricultural Sciences, 298(12): 1401-6230. |
Wilson B F, Archer R R. 1977. Reaction wood: induction and mechanical action[J]. Annual Review of Plant Physiology, 28(6): 23-43. |
Yamaguchi K, Shimaji K, Itoh T. 1983. Simultaneous Inhibition and Induction of compression wood formation by morphactin in artificially inclined stems of japanese larch (Larix leptolepis Gordon)[J]. Wood Sci Technology, 17(2): 81-89. DOI:10.1007/BF00369125 |
Yamashida S, Yashida M, Yamamoto H. 2009. Relationship between development of compression wood and gene expression[J]. Plant Science, 176(6): 729-735. DOI:10.1016/j.plantsci.2009.02.017 |
Yoshizawa N, Idei T. 1987. Some structural and evolutionary aspects of compression wood tracheids[J]. Wood Fiber Sci, 19(4): 343-352. |
Yumoto M, Ishida S. 1983. Studies on the formation and structure of compression wood cells induced by artificial inclination in young trees of Picea glauca: Ⅲ. Light microscopic observation on the compression wood cells formed under five different angular displancements[J]. Journal of the Faculty of Agriculture, Hokkaido University, 60(4): 337-351. |
Zobel B, Van Buijtenen J P. 1989. Wood variation-its causes and controls[M]. Berlin: Springer Verlag.
|