 2012, Vol. 48
2012, Vol. 48文章信息
- 邓顺, 彭观地, 舒金平, 王浩杰
- Deng Shun, Peng Guandi, Shu Jinping, Wang Haojie
- 居竹伪角蚜种群的年动态变化及调节因子
- Investigation of Year-Round Population Dynamics of Pseudoregma bambusicola and Its Regulating Factors
- 林业科学, 2012, 48(1): 103-108.
- Scientia Silvae Sinicae, 2012, 48(1): 103-108.
-
文章历史
- 收稿日期:2010-09-10
- 修回日期:2011-10-17
-
作者相关文章



2. 江西省赣州市林木种苗站 赣州 341000
2. Forest Seed and Seedling Station of Ganzhou Ganzhou 341000
居竹伪角蚜(Pseudoregma bambusicola)又名竹茎扁蚜(以下简称竹蚜), 属半翅目(Hemiptera)扁蚜科(Hormaphididae)伪角蚜属(Pseudoregma), 主要分布于日本、中国、泰国、印度等亚洲东、南部地区, 在国内主要分布于浙江、福建、湖南、江西、四川、广东、广西、台湾等亚热带地区(Fukatsu et al., 2001; 徐天森等, 2004; 郭美华等, 2006)。居竹伪角蚜主要危害孝顺竹(Bambusa multiplex)、观音竹(B.multiplex cv.Fernleaf)等丛生类竹种, 受害寄主均为1年生新竹, 蚜虫群聚于竹杆表面或嫩枝上取食, 危害严重时遍布整个竹杆, 成蚜取食时在节间分泌蜜露, 使竹叶和枝条上感染煤污病, 新竹存活率严重下降。对居竹伪角蚜的生物学特征、化学防治措施、COI遗传型鉴定、种群空间分布及其天敌种类等已有研究(Sakata et al., 1991; 易龙, 2003; 于炜等, 2006; 见许文馨, 2007; 邓顺等, 2009), 但迄今还未见种群年动态变化及外界生态因子对其影响方面的研究。由于此类蚜虫多危害观赏性竹种, 单一喷施化学药剂可能对旅游资源及生态环境造成污染, 因此, 本文在前期调查的基础上, 结合主要天敌、气候(降雨)等生态因子, 进一步研究居竹伪角蚜种群的年动态变化, 旨在发现对其种群有关键性制约的调节因子, 从而提出高效、便捷的生态防治方法, 能在短时间内有效控制该蚜虫的大规模发生。
1 材料与方法 1.1 样地概况所选样地位于浙江省富阳市大桥路73号的中国林业科学研究院亚热带林业研究所竹林小道(30°16' 43″N, 120°12'01″E), 样地种植有年限>10年的孝顺竹种群, 共包括4块样地, 分别以Ⅰ、Ⅱ、Ⅲ和Ⅳ标号表示, 各样地间隔>10 m, 各样地内竹丛数不同(样地Ⅰ22丛, 样地Ⅱ8丛, 样地Ⅲ24丛, 样地Ⅳ7丛), 竹丛之间为2~3 m的间隔, 竹笋出土时间为5月底至7月中旬。样地植株处于自然生长状态, 无人为干扰, 生长区域为典型的亚热带气候特征。
1.2 竹蚜种群密度及其受害率的调查竹蚜种群动态的调查方法分为2种:1)密度调查从2008年7月初至2009年5月初, 从4块样地中随机选择竹径基本一致(21~25 mm)的受害新竹15~20株作为样株, 每次调查时1株只取1次样, 调查时间间隔10~15天, 每月调查2~3次, 对受害植株依次编号, 以避免连续2次对同一植株取样; 调查过程中若选定的样株死亡或恢复健康水平, 则另外选取邻近受害株替代, 以保证每次能获得较为稳定的竹蚜种团。取样部位为受害株中部(离地2~3 m), 采用黄板粘贴法粘取受害株上的蚜块(邓顺等, 2009), 带回实验室后统计黄板上粘有的竹蚜总数, 分别统计出1~2龄和3~4龄的蚜虫数, 1~2龄中包括兵蚜数; 2)危害率调查依次统计样地中各竹丛的危害株数, 统计每月所有样地的新竹受害株数, 同时转换为各样地新生竹的受害百分率。
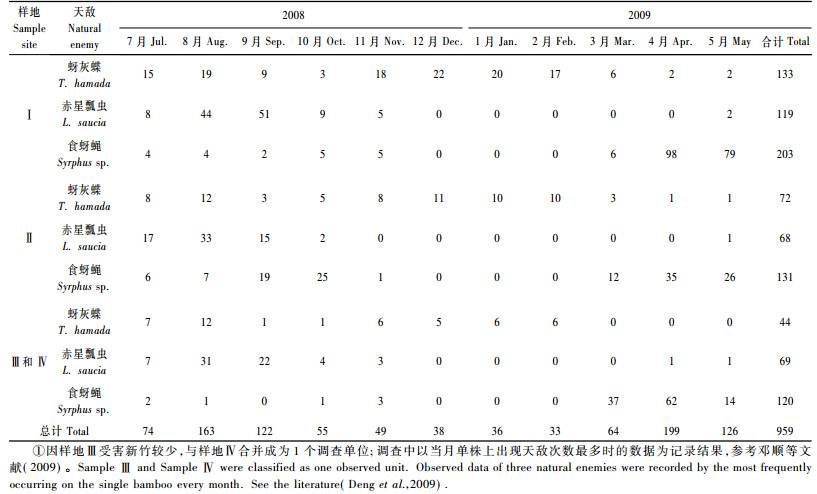
1.3 竹蚜天敌种群密度调查根据编号依次调查竹丛中受害单株上的天敌种类及其数量, 与危害率调查同步进行, 统计方法参考邓顺等(2009), 即以当月单株上出现天敌次数最多时的数据作为记录结果。捕食竹蚜的3种天敌中仅赤星瓢虫(Lemnia saucia)在幼虫和成虫阶段均能取食竹蚜, 其他2种天敌仅以幼期虫态取食, 记录时不包括蚜灰蝶(Taraka hamada)和食蚜蝇(Syrphus sp.)的成虫在内。
1.4 竹蚜迁移习性观察从2008年5月底开始, 观察新竹上的竹蚜发生动态, 记录若蚜在竹丛中扩展情况。选取样地Ⅰ中的4个竹丛, 每隔5~7天观察1次, 每次60 min, 在每天10:00—11:00之间进行, 此时若蚜活动能力较其他时间段强, 同时用记号笔在受害株上标记。从10月末开始观察受害株上是否出现有翅蚜, 并通过黄板粘贴法计算方块内有翅蚜的密度, 同时在受害株附近的新竹和杂草中布置剪切好的黄板(10 cm × 10 cm), 以检测有翅侨蚜的迁飞状况。
1.5 统计分析利用独立样本t测验检验全年不同时期竹蚜种群密度的稳定性。
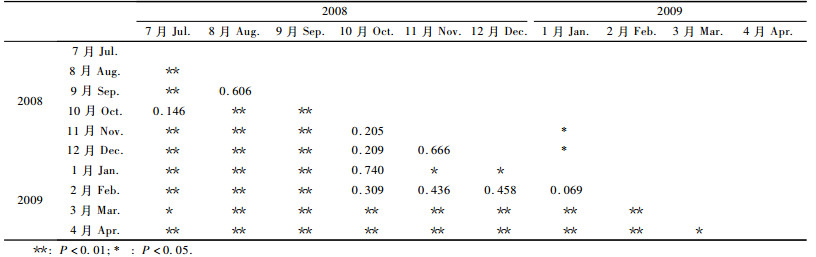
2 结果与分析 2.1 不同月份的竹蚜种群密度对调查数据进行独立样本t测验, 结果表明(表 1):2008年8—9月、2009年3—4月竹蚜种群平均密度出现2个高峰期, 均与其余月份种群平均密度之间差异达到极显著(P<0.01); 2个高峰期之间竹蚜种群平均密度差异也极显著(P<0.01), 8—9月种群密度的平均值高于3—4月。图 1显示了1年中竹蚜种群平均密度的动态, 也显示了2008年8—9月和2009年3—4月的2个竹蚜种群密度高峰期。
|
|

|
图 1 2008-07—2009-05不同月份竹蚜种群的平均密度 Figure 1 Mean value of population density of P.bambusicola in different months from July in 2008 to May in 2009 |
从2008年的10月初至2009年3月中旬, 竹蚜种群龄期比例处于较高的阶段(图 2), 平均值在0.792~1.250之间, 总平均值为0.912 ± 0.117, 表明这段时期3~4龄竹蚜数量较多, 干母产子率不高; 而其余月份竹蚜种群的龄期比例较低, 平均值在0.5上下(0.392~0.667), 其中2008年7月初至9月末的总平均值为0.510 ± 0.075, 2009年3月末至5月初的总平均值为0.478 ± 0.086, 表明在这2个时间段中若蚜出生率较高, 其数量与3~4龄蚜虫数量相当。

|
图 2 2008-07—2009-05不同月份竹蚜种群龄期段的比例 Figure 2 Age ratio between the 3rd~4th instar and the 1st~2nd instar in different months from July in 2008 to May in 2009 |
在各样地中, 3种天敌中以食蚜蝇的发生数量最大(表 2), 赤星瓢虫发生高峰期在新竹发生的2008年7—9月, 食蚜蝇发生高峰期在2009年的3—5月, 而蚜灰蝶发生高峰期在全年中有2个, 分别为2008年的7—8月和11月—2009年2月(图 3)。3种天敌中仅蚜灰蝶全年伴随竹蚜存在, 赤星瓢虫和食蝇蝶则在气温较低时期(12月中旬至2月底)以成虫藏匿于竹子箨叶或竹丛周围的杂草中越冬, 3月初温度回升时才出来活动。
|
|

|
图 3 2008-07—2009-05不同月份间样地内3种天敌的数量 Figure 3 Total number of three natural enemies in different months from July in 2008 to May in 2009 |
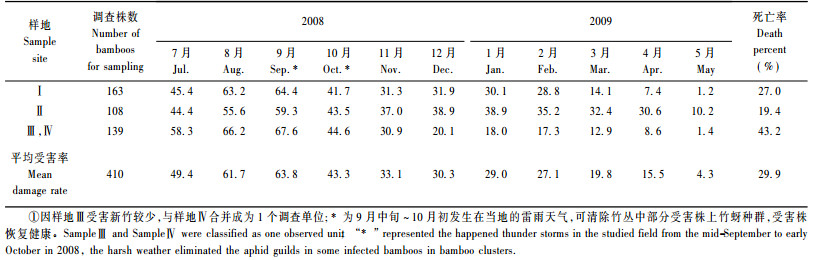
从表 3可以看出, 2008年8—9月为新竹受害高峰期, 从10月开始逐渐下降, 一直到2009年5月份, 受害株数趋近于0, 从2009年3月初开始, 各竹丛中陆续有受害植株死亡, 死亡率在19.4% ~ 43.2%之间。综合表 2和表 3的调查结果可以发现:竹丛受害株数的减少与3种天敌的发生有一定的关联, 2008年竹蚜密度发生高峰期在8—9月, 之后陡然下降, 这一时期赤星瓢虫的大发生可能是导致新竹受害率降低的因素之一, 2009年4—5月受害率进一步降低, 趋近于零, 可能与这一时期食蚜蝇的大发生有着密切的关系; 在2008-11—2009-02之间, 蚜灰蝶发生数量在3种天敌中占优, 但由于各天敌种群密度过低, 各竹丛受害率并未有显著的下降。此外, 9—10月中连续数天的雷雨天气也可清除受害植株上的竹蚜种群。
|
|
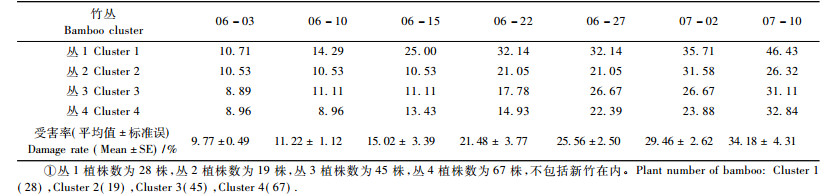
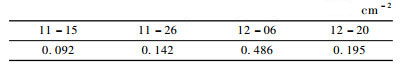
新竹出土后即被竹蚜危害, 初期时1龄若蚜数量较多, 多聚于新竹基部取食, 由于竹丛中新竹相隔很近, 若蚜很快向邻近新株扩散, 从表 4可以看出, 样地Ⅰ各竹丛受害率发展迅速, 呈现典型的由点到片的危害模式。有翅侨蚜出现时间1个月左右, 从11月中旬开始, 12月初达到密度高峰期(表 5), 一直到12月末结束。有翅侨蚜发育完全后, 一些迁移至未被竹蚜危害的植株, 并在其基部产下若蚜, 但若蚜存活几率不高, 受害株很快能恢复健康, 试验中也观察到有翅侨蚜能迁飞至竹丛四周的杂草中。
|
|
|
|
以往研究表明:危害观音竹的居竹伪角蚜在全年存在2个发生高峰期, 分别为5—6月和9—11月(于炜等, 2006), 本试验也表明该竹蚜在全年中有2个发生高峰期, 但发生期与前者不同, 分别为2008年的8—9月和2009年的3—4月, 这可能与寄主种类、竹笋出土时期以及竹蚜所处的地理环境相关, 如观音竹属于孝顺竹的一个变种, 在出笋时间上, 杭州植物园孝顺竹出笋时间为每年的3—5月, 而富阳地区的孝顺竹出笋时间为每年的5—7月(朱石麟等, 1994; 于炜等, 2006)。2008年8—9月竹蚜种群的平均密度高峰期可能与新竹生长时积聚起来的营养水平密切相关, 新竹在生长过程中不断积聚土壤中的养分, 为竹蚜种群的发展提供适宜的生境和营养组分, 而2009年的3—4月为资源的枯竭期, 这段时间竹丛中的不少受害株死亡, 此时还未有新竹出现, 竹蚜种群被迫往其他营养未耗尽的受害株迁移, 暂时性地造成单位种群密度的增加, 导致在新竹更替之前的第2个密度高峰期的出现, 这个高峰期形成的原因也与以往报道的3—4月的高峰期不同(易龙, 2003)。
在全年过程中, 1~4龄若蚜均同时发生, 不同龄期若蚜密度的比例反应了不同时期竹蚜通过调整种群自身的年龄组配来适应寄主的营养状况或环境条件。由于居竹伪角蚜种群密度大, 1龄若蚜与2龄若蚜、3龄竹蚜与4龄竹蚜在形态和大小上非常接近, 在实际调查过中难以将1~2龄若蚜、3~4龄若蚜区分开来, 但3~4龄与1~2龄竹蚜间的形态差别比较明显, 因而本试验调查时分别以1~2龄和3~4龄分组进行统计。在新竹发生期间(2008-10—2009-02), 3~4龄若蚜的比例平均值为0.912 ± 0.117, 表明若蚜出生率不高, 种群中以3~4龄的蚜虫为主, 这种情况的发生可能与较低的气温有关; 其他月份该比例的平均值在0.5上下, 1~2龄竹蚜的数量与3~4龄竹蚜的数量大致相当, 各龄期蚜虫都有均等的机会利用新竹的营养成分, 同时与合适的温度有一定关联。
居竹伪角蚜3种主要天敌的发生时间具有一定规律性。在新竹受害率与不同天敌之间, 赤星瓢虫和食蚜蝇发生的高峰期新竹受害率明显降低。赤星瓢虫发生高峰期为8—9月, 这段时间竹蚜发生严重, 新竹受害率为55.6%~67.6%, 为全年受害最重的时期, 10月份受害率降至45.0%以下, 表明这段时期竹蚜种群可能受到瓢虫的抑制。3—5月食蚜蝇的大量发生伴随着新竹受害率的骤减, 但同一时期除食蚜蝇的影响外, 受害株死亡或营养水平的下降也可能是导致新竹受害率降低的影响因素之一, 有些竹丛新竹的死亡率最高可达40.0%以上。此外, 发生在9—10月的雷雨天气, 可清除竹丛中部分受害较轻的植株上的蚜块。在资源紧缺的情况下居竹伪角蚜分别通过若蚜和有翅侨蚜扩散到健康新竹上, 发生时间分别是新笋出土时期和全年气温较低的季节, 若蚜的扩散能力可能比有翅侨蚜有效, 在有翅蚜出现的月份中, 受害植株发生率并未增多, 而在新竹刚出土的时期, 若蚜扩散速度较快, 致新竹受害率迅速增加。
上述试验观察表明:在新竹即将完成第1年生长的最后时期以及下一年新竹出土时, 是竹蚜滋生蔓延的较好时机, 此时竹丛中新笋出土数量不多, 如表 4所示, 在这一时期若采用合适的防治方法或能有效地抑制竹蚜的普遍发生。本研究中只考察了3种天敌对竹蚜滋生的影响, 也初步证实了赤星瓢虫和食蚜蝇可能是制约蚜块扩散的重要因子, 有研究通过捕食性功能反应表明蚜灰蝶对另一种竹蚜—竹叶扁蚜(Astegopteryx bambusifoliae)种群有显著的控制作用, 是该竹蚜的优势天敌种类(丁玉洲等, 2010), 但本试验通过田间调查却发现蚜灰蝶对于居竹伪角蚜种群的扩散影响较小, 因此, 有必要结合3种天敌的捕食功能性反应对制约蚜块的关键因子作出进一步的分析。
邓顺, 王鹏, 舒金平, 等. 2009. 居竹伪角蚜及其3种主要天敌的空间格局研究[J]. 环境昆虫学报, 31(4): 300-305. |
丁玉洲, 许明修, 刘小林, 等. 2010. 蚜灰蝶对竹伪蚜的捕食效应[J]. 林业科学, 46(6): 93-96. DOI:10.11707/j.1001-7488.20100614 |
郭美华, 谢易霖, 丘明智. 2006. 日月潭地区陆域昆虫多样性调查[J]. 台湾昆虫, 26(2): 125-142. |
徐天森, 王浩杰. 2004. 中国竹子主要病虫害[M]. 北京: 中国林业出版社.
|
许文馨. 2007. 竹茎扁蚜之群落混合. 台北: 台湾大学昆虫学研究所硕士学位论文.
|
易龙. 2003. 10%吡虫啉对居竹伪角蚜的防治试验[J]. 植物医生, 16(3): 25-26. |
于炜, 曾新宇, 徐芸茜, 等. 2006. 竹茎扁蚜生物学特性与防治[J]. 湖北植保, 13(5): 12-13. |
朱石麟, 马乃训, 傅懋毅. 1994. 中国竹类植物图志[M]. 北京: 中国林业出版社.
|
Fukatsu T, Shibao H, Nikoh N, et al. 2001. Genetically distinct populations in an Asian soldier-producing aphid, Pseudoregma bambucicola(Homoptera:Aphididae), identified by DNA fingerprinting and molecular phylogenetic analysis[J]. Molecular Phylogenetics and Evolution, 18(3): 423-433. DOI:10.1006/mpev.2000.0887 |
Sakata K, Itô Y. 1991. Life history characteristics and behaviour of the bamboo aphid, Pseudoregma bambucicola(Hemiptera:Pemphigidae), having sterile soldiers[J]. Insects Sociaux, 38(3): 317-326. DOI:10.1007/BF01314917 |