 2012, Vol. 48
2012, Vol. 48文章信息
- 王文杰, 孙伟, 邱岭, 祖元刚, 刘伟
- Wang Wenjie, Sun Wei, Qiu Ling, Zu Yuangang, Liu Wei
- 不同时间尺度下兴安落叶松树干液流密度与环境因子的关系
- Relations Between Stem Sap Flow Density of Larix gmelinii and Environmental Factors under Different Temporal Scale
- 林业科学, 2012, 48(1): 77-85.
- Scientia Silvae Sinicae, 2012, 48(1): 77-85.
-
文章历史
- 收稿日期:2011-01-07
- 修回日期:2011-03-25
-
作者相关文章



全球变化过程如何影响水分循环, 一直是全球变化生态学研究的重点领域之一(方精云, 2000)。森林生态系统在调节水分循环和小气候方面具有重要功能, 其基础之一是树木个体对水分运转的影响。对森林水分循环研究的基础是了解树木个体如何对外界环境因子的响应, 如能对其准确客观描述, 将有助于了解树木个体、乃至森林生态系统在全球变暖的进程中的响应。对树木个体水分循环研究的一个重要方法是基于热扩散技术的树干液流观测(Swanson et al., 1981; Anfodillo et al., 1993; Swanson, 1994; Lindroth et al., 1995; Granier, 1987; 王华田, 2002; 王瑞辉等, 2006), 前人在这方面已进行不少研究, 如:刘奉觉等(1993)对杨树(Populus)树干液流时空动态研究; 孙鹏森等(2000)对油松(Pinus tabulaeformis)树干液流的时空变异性研究; 李海涛等(1998)应用热脉冲技术对棘皮桦(Betula luminifera)和五角枫(Acer frictum)树干液流的研究。这些对了解不同树种树干液流特点具有重要功能。在环境因子如土壤和空气温度、土壤和空气湿度、光照等与树干液流关系的研究方面, 因需长期环境因子与树干液流等同时连续测定, 对设备及投入要求较高而较少进行(尹光彩等, 2004; 马玲等, 2005; 王翠等, 2008)。本文以我国东北地区重要造林树种兴安落叶松(Larix gmelinii)为对象, 连续1年观测, 旨在分析:1)环境因子对树干液流影响的显著性是否存在时间尺度差异; 2)这种不同时间尺度下环境因素影响树干液流的模式对简化分析树干液流特征及耗水总量估计是否具有意义; 3)单株落叶松是否能代替这个林分其他落叶松表达对环境因子的响应。
1 研究地概况研究地位于东北林业大学帽儿山试验林场老山试验站(45°20' N, 127°34' E)。属长白山系张广才岭西北部余脉, 平均海拔约340 m。气候属温带大陆性季风气候, 四季分明, 冬长夏短, 雨量集中在7—8月。年平均气温2.8 ℃, 年平均相对湿度70%左右, 年平均降雨量723.8 mm, 年平均蒸发量1 093.9 mm, 年平均积温2 526 ℃, 无霜期120~140天, 年日照时数为2 471.3 h, 2003—2005年的平均降雨量为650 mm。观测站位于试验站林场西北部的落叶松人工林内。坡向南, 坡度5°左右, 郁闭度0.7左右。主要树种为兴安落叶松, 1969年造林, 平均株高22 m左右, 胸径20 cm左右。
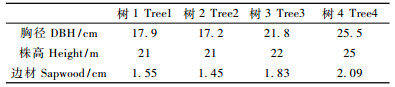
2 材料与方法 2.1 试验材料在人工林内选择4株落叶松进行树干液流测定, 4株落叶松的生长状况如表 1。
|
|
利用热扩散探针产生的温差来计算树干液流通量密度, 在高约1.4~1.6 m的树干处, 用刀将树干死皮刮掉, 不能损坏树木形成层。用电钻打4 cm左右深、直径为3 mm的2个圆孔, 其距离为10 cm, 然后把TDP热扩散探针(Thermal Dissipation Probe, Dynamax, 美国)安装进去, 用橡皮泥封好, 避免雨水进入以造成监测结果误差过大和仪器损伤。外面用防护罩罩上, 防止鸟类和自然造成的机械损伤。本套设备的热探头由2根探针组成, 上部的探针能恒定持续加热, 内部含有热元件和热电偶; 下部探针为参考端, 只有热电偶。设置每30 min采集1次平均数据, 时间为2005年1月1日—12月31日。
瞬时树干液流密度的计算公式为:
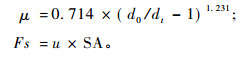
|
式中:d0, 无液流时的温差值(℃); dt, 当时测定的温差值(℃); μ的单位为mL·cm-2 min-1; SA, 横断面边材面积(cm2)。
2005年1月1日—12月31日环境因子数据采用Campbell CR10X数据采集器同步采集。光照(ML-020P, EKO, 日本)、空气温度和空气湿度(HMP-45D, Vaisala, 芬兰)均在23 m高度采集, 土壤温度(C-PTWP-10, Climatec, 日本)和土壤湿度(CS615, Campbell, 美国)均在5 cm深采集。由于供电系统和雨季打雷对仪器的破坏等原因, 导致如下数据缺损:液流密度缺少第9, 16, 84, 153, 168, 212到224, 255, 310~354天(日历日, Calendar day)的数据; 空气温度缺少118~147, 152~185及215天的数据; 光照和空气湿度缺少152~174及215天的数据; 土壤温度和土壤湿度缺少26~31天的数据。
2.2.2 数据分析因为停电或其他原因会产生过大或过小、不合乎常理的异常值, 需要在分析之前检测找出剔除, 以保证对研究结果不产生影响。本文运用以下方法来判断和分析数据质量, 以便进行数据质量控制:1)通过对数据作图, 从直观角度对异常数据进行判断; 2)当数值超出所测定区间平均值± 3SD的情况下, 认为其值异常, 去除平均值用相邻数值的平均值替代。
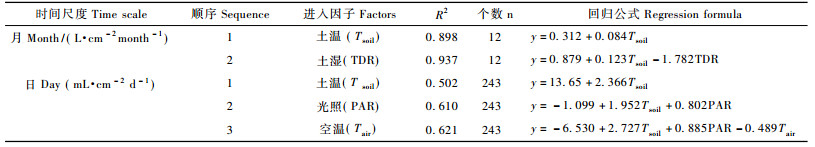
为了探讨不同时间尺度条件下树干液流与环境因子的相关关系, 首先把树干液流密度以及相关的环境因子转换为小时、日、月尺度相应数值。小时尺度数据为各仪器所测定的指标, 测定频度为半小时; 日尺度数据中土壤温度、空气湿度、土壤湿度和空气温度取每天测定所有数据(每天48个)的平均值, 而光照和树干液流密度通过时间积分方法, 由原单位μmol·m-2 s-1和mL·cm-2 min-1转换为moL·m-2 d-1和mL·cm-2 d-1; 月尺度数据中中土壤温度、空气湿度、土壤湿度和空气温度取每天平均值(每月28~31个)的平均值, 而光照和树干液流密度通过时间积分方法, 由原单位mol·m-2 d-1和mL·cm-2 d-1转换为mol·m-2 month-1和mL·cm-2 month-1。
按照逐步进入的方式, 将月尺度和日尺度下环境因子与树干液流关系进行逐步回归分析。在小时尺度上, 环境因子对树干液流密度存在时滞影响(谢恒星等, 2007; 王华等, 2009; 孙迪等, 2010), 通过错位分析的方法, 确定不同季节、不同环境因子对树干液流的时滞效应, 并基于这一事件对原始数据进行校正。对经过时滞校正的数据, 同样按照逐步进入的方式, 将小时尺度下环境因子与树干液流关系进行逐步回归分析, 各因子对树干液流的影响从进入顺序、斜率大小和相关系数R2来判断。
具体原则:进入越早, 则说明其影响越大, 对于月、日尺度条件下, 因为只有一次逐步回归分析, 容易判断。对于小时尺度上, 由于是分季节进行, 所以会出现不同的进入次序, 为了判断其影响大小, 计算各因子在不同季节逐步回归进入顺序之和, 并依据进入顺序之和的大小, 对不同因子的影响进行判断, 和越小, 说明更多的情况其对树干液流的影响较大; 反之, 则影响较小。斜率大小反映了单位环境因子变化所导致树干液流变化的大小, 取各次逐步回归中各因子相应斜率进行平均处理, 代表这一因子变化所导致树干液流的变化大小。有些因子在某些逐步回归中没有进入, 给其赋值为6(因为5个因子, 各因子进入顺序最大为5)。决定系数R2的生态学意义是现有拟合方程能够解释因变量产生变异的百分率, 其值约接近于1(100%), 说明所选环境因子的变化越能够更好地解释因变量期间产生的变化。本文所有统计分析均使用SPSS17.0进行线性逐步回归分析。
3 结果分析 3.1 月尺度下树干液流与环境因子相关性分析树干液流主要集中在5—10月之间, 占全年总量的85%, 其中8月树干液流密度平均值最高达到2 L·cm-2 month-1以上, 6, 7和9月在1.5 ~ 1.8 L·cm-2 month-1之间。树干液流密度月平均值的最低值出现在11和12月, 其中最低的11月为0.055 L·cm-2 month-1(图 1)。

|
图 1 树干液流密度与气温、土温、光照、空气湿度、土壤湿度的月均值变化 Figure 1 Changes of monthly sap flow density, air temperature(Tair), photosynthetic active radiation(PAR), air relative humidity(RH), soil temperature(T soil), and soil humidity measured by time domain reflectometer(TDR) |
空气温度的月均值呈现1个单峰曲线, 7月最高温为20.8 ℃, 12月最低温达到-17.88 ℃。光照则呈现出双峰曲线, 峰值在3和8月, 低值出现在1, 6和12月, 其中8月最高值达到890 mol·m-2 month-1。空气湿度则多在50%~90%之间, 其中7月峰值为88.4%, 低值分别在3月和10月(50%)。土壤温度和土壤湿度都是1个单峰变化的曲线, 最高值出现在7—8月(土壤温度:18 ℃; 土壤湿度:0.72), 低值出现在12月—次年2月(土壤温度:-3 ℃; 土壤湿度:0.21)(图 1)。
逐步回归分析结果显示:土壤温度对树干液流的影响最大, 可以解释树干液流变化的89.8%, 土壤温度升高1°树干液流可以提高0.084 L·cm-2 month-1; 第2进入的因子为土壤湿度, 二者可以解释93.5%的树干液流月尺度上的变化, 与土壤温度不同, 土壤湿度与树干液流密度呈负相关(表 2)。土壤温度和土壤湿度单位增加分别引起树干液流上升0.084~0.123 L·cm-2 month-1和下降1.782 L·cm-2month-1(表 2)。
|
|
总体上来说, 树干液流密度日均值在130~300天比较大, 约为20~90 mL·cm-2 d-1, 平均值为50 mL·cm-2 d-1; 但是在70~90天之间也出现1个峰值, 约为50 mL·cm-2 d-1(图 2)。

|
图 2 树干液流密度与气温、土温、光照、空气湿度、土壤湿度的日均值变化 Figure 2 Changes of daily sap flow density, Tair, PAR, RH, T soil, and TDR |
空气温度日均值则是单峰曲线, 最大值出现在221~227天之间, 日均温度为25.6~25.8 ℃, 最小值约为-23.4 ℃, 出现在351天; 光照最高出现在180天左右, 高达50~60 mol·m-2 d-1, 冬季较低, 多在5~15 mol·m-2 d-1之间。空气湿度多在30% ~ 95%之间, 不同季节间的变化并不明显。土壤温度和土壤湿度均为单峰变化, 其中土壤温度最大值约22 ℃, 最小值约-4 ℃, 土壤湿度最大值在0.80, 最小值为0.17左右(图 2)。
逐步回归分解结果表明:土壤温度对树干液流影响最大, 能够解释其变化的50.2%, 土壤温变每升高1°导致树干液流密度增强2.366 mL·cm-2 d-1, 光照是第2顺次引入的因子, 相关系数R2提高到0.610, 其与树干液流存在正相关, 第3引入因子是空气温度, R2提高到0.621。土壤温度为最主要的影响因子, 单位增加导致树干液流上升1.9~2.7 mL·cm-2 d-1; 光照单位提高可以导致树干液流0.8~0.9 mL·cm-2 d-1的升高, 而空气温度单位升高将引起0.489 mL·cm-2 d-1的降低(表 2)。
3.3 小时尺度下树干液流密度和环境因子相关性分析及其季节差异在春季(以2005年第70~72天为例)、夏季(以2005年第187~189天为例)、秋季(以2005年第256 ~ 258天为例)和冬季(以2005年第356~358天为例)选择3天数据比较环境因子和树干液流密度日变化差异(图 3)。空气温度昼夜变化最大的是秋季, 夜间温度多在10 ℃左右, 而午间温度可以达到21 ℃左右, 夏季夜间温度多在16°左右, 午间温度在22~25 ℃之间, 多出现在下午14: 00—15: 00之间, 冬季和春季同样存在明显的空气温度昼夜变化。空气湿度在春季和秋季昼夜变化最大, 其次是夏季, 冬季变化最小, 一般是14: 00—15: 00空气湿度降低到最低点, 夜间空气湿度最高, 不同季节多在80%以上, 夏季甚至达到100%。光照夏季最强, 最高可以到7~8 mol·m-2 h-1, 其次是秋季和春季, 最高多在4~6 mol·m-2 h-1。冬季最高多在1~2 mol·m-2 h-1之间。冬季和春季土壤温度日变化不明显, 夏季类似, 最为明显的是秋季, 昼夜温差可以达到2°以上。土壤湿度秋季、春季和冬季均没有明显昼夜变化, 而夏季主要是由于降雨的发生(189天下雨), 可以引起土壤湿度显著变化0.2(v/v)以上(图 3)。

|
图 3 不同季节树干液流密度与气温、土温、光照、空气湿度、土壤湿度的日变化差异 Figure 3 Changes of hourly sap flow density, Tair, PAR, RH, T soil, and TDR. |
树干液流密度显著昼夜变化发生在秋季和夏季, 最大值多出现在9 : 00—11 : 00点间, 夏季4 ~ 5 mL·cm-2 h-1, 而秋季可以达到7~8 mL·cm-2 h-1, 冬季和春季液流密度较小, 昼夜变化不大:冬季12: 00左右达到峰值0.2~0.4 mL·cm-2 h-1左右, 春季出现时间相差不大, 峰值在0.7~1.1 mL·cm-2 h-1, 在晚上和凌晨达到最低值, 趋于0(图 3)。
这种环境因子不同季节间的差异, 对树干液流的影响, 本文通过逐步回归分析方法进行了分析(表 3)。对春季(61~151天)树干液流密度与5个环境因子进行逐步回归分析, 依次入选的环境因子为空气湿度、土壤温度、空气温度和光照, 其中空气湿度能够解释2 937组数据变异的16%, 土壤温度和空气湿度共同解释其中27%的变化, 4个环境因子能够解释其中31%的变异。夏季(152~243天), 空气湿度能够解释树干液流变化的61%, 其后顺次进入空气温度、光照、土壤温度和土壤湿度, 这些因子共同能够解释树干液流76%变异。对秋季(244~304天)数据, 依次进入光照、空气温度、空气湿度和土壤湿度, 4个因子能够解释树干液流发生变化的45%, 而第1进入因子光照可以解释32%。对于冬季(1~60, 305~365天), 影响树干液流密度的主次顺序为光照、土壤湿度、空气温度、空气湿度, 能够解释树干液流发生变化的39%, 同样第1进入因子光照解释32%。公开全年数据, 光照对树干液流密度影响最大, 其次是土壤温度、空气湿度和空气温度。总体来看, 环境因子与树干液流相关程度存在季节性差异:夏季的相关性最高, 在0.614~0.758之间; 春季相关性最低, 仅0.162~0.306之间; 其他秋季和冬季多在0.3~0.4之间; 全年的数据相关性在0.2~0.4之间。分析外界环境因子对树干液流的影响, 最佳的季节应在夏季:这一季节树干液流发生变化的60%~75%可以由外界环境变化来解释。
|
|
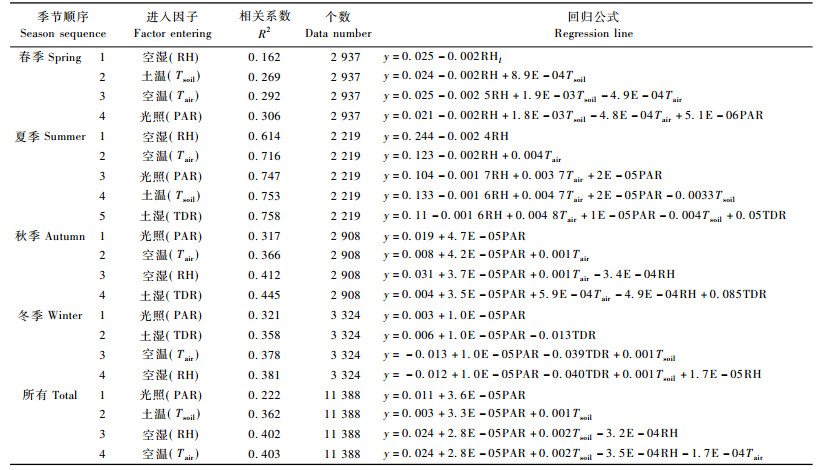
树干液流密度与外界环境因子线性相关的斜率具有生物学意义, 其绝对值大小反映了所测定指标对树干液流密度的影响程度, 而其值正负则反映了影响方向。表 4是逐步回归分析中同一环境因子的斜率进行的统计分析, 空气湿度对树干液流的影响多数季节均表现为负相关, 即湿度越高, 树干液流越小。冬季虽然为正相关, 但是其绝对值大小仅为其他季节的1/20~1/100。空气湿度每增加1个单位, 树干液流下降最高发生在春季, 达到0.127 5 mL·cm-2 h-1, 最低发生在冬季, 稍微上升0.00 1 mL·cm-2 h-1, 全年平均下降0.056 4 mL·cm-2 h-1。土壤温度对树干液流影响不同季节不一致, 秋季和冬季没有显著影响, 而春季为正影响, 夏季则为负影响, 全年数据分析表现为正影响, 平均值表现为很小的负影响(-0.009 1)。空气温度对树干液流的影响多表现为正的影响, 如夏季、秋季和冬季, 但是当把全年数据一起统计时, 表现为负影响, 这是受春季数据影响的结果, 平均值表现为正的影响, 空气温度每升高1 ℃, 能够引起树干液流密度增加0.066 1个单位。光照对树干液流的影响在不同季节均表现正向影响, 夏季和秋季光强增加1 mmol·m-2s-1树干液流能够增加1~2.4 mL·cm-2 h-1, 而春季和秋季则较小, 只有0.3~0.6 mL · cm-2 h-1, 平均值为1.24 mL·cm-2 h-1。全年数据分析和春季数据表现为土壤湿度对树干液流影响不大, 而夏季、秋季和冬季数据也表现出不同的影响:夏季和秋季正向影响, 而冬季为负向影响, 平均值表明:土壤湿度增加1个单位, 树干液流增加2.1 mL·cm-2 h-1。
|
|
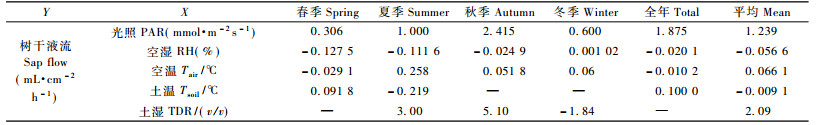
逐步回归中因子进入得越早, 说明其对树干液流影响可能越大。为了分析各个因子对树干液流影响的强弱, 笔者把全年数据逐步回归分析中各因子进入顺序相加, 作为评价各因子的重要性的一个指标。各因子所得越小, 说明其逐步回归进入得越早, 即对树干液流影响越大。有些因子在某些逐步回归中没有进入, 笔者将其赋值为6(因为5个因子, 各因子进入顺序最大为5)。由图 4可以看出:在小时尺度上, 对树干液流影响最大的是光照, 其次是空气湿度、空气温度, 而土壤温度和土壤湿度影响最小。

|
图 4 基于逐步回归进入顺序之和比较小时尺度上不同因子对树干液流影响的重要性 Figure 4 Comparison of different environmental factors influences on stem sap flow according to the sum of entering sequences in stepwise regression analysis |
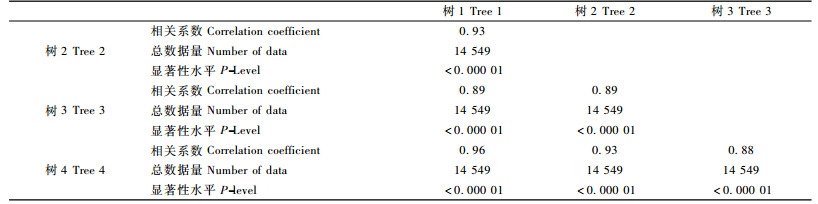
对4株样树树干液流进行相关分析如表 5所示。14 549个数据分析得到相关系数分别为0.93, 0.89, 0.96, 0.89, 0.93, 0.88, 且P-level值均小于0.000 01, 说明多数情况下, 树干液流不同个体之间差异不大, 具有高度的一致性。
|
|
树干液流的产生主要是由于蒸腾拉力作用(Ford et al., 2007), 这些水分用于叶片和不同组织的生化反应(如光合呼吸)以及保持植株个体状态。此外, 影响树干液流的因素还包括物理环境因素, 如空气湿度与植物组织器官内湿度差所导致的直接蒸发作用、土壤湿度与根系之间的水分差异等(Nicolas et al., 2005; Becker, 1998)。从理论上讲, 能够影响这些过程的环境因子, 如研究所选的光照、空气温度、土壤温度、空气湿度、土壤湿度均能够直接或者间接影响树干液流特征。
以往研究环境因子影响树干液流密度变化结论不尽一致, 如Granier等(1996; 1998)对21年生挪威云杉(Picea abies)和热带雨林研究发现:空气湿度对树干液流的影响较光照或空气温度显著; 熊伟等(2003)认为太阳辐射、空气温度和空气相对湿度是影响单木蒸腾量的主要气象因子; 尹光彩等(2003)认为影响桉树(Eucalyptus)人工林液流通量的主要环境因子是空气水气压差和土壤水分。在不同时间尺度上对树干液流与环境因子相关关系的研究, 能够帮助人们从树干液流的机制过程角度理解如何准确估计树干耗水总量; 在何种尺度下, 估计树干液流应该需要考虑哪些环境因子。目前, 在大尺度(月或者年)上, 树干液流研究很多集中在林分耗水量及其季节变化上(王华田等, 2002; 王瑞辉等, 2006), 而在较小尺度上(小时)多是影响树干液流的时滞效应进行分析及其生理生态学意义(谢恒星等, 2007; 王华等, 2009; 孙迪等, 2010)。通过对落叶松成树1年的研究, 建立了不同时间尺度(月、天、小时)上环境因子与树干液流密度的相关关系, 并进行逐步回归分析, 旨在发现时间尺度对树干液流与环境因子关系的影响模式以及其对林分耗水研究的意义。
随着时间尺度的减小, 光照对树干液流的影响逐渐增大, 而土壤温度和湿度的影响逐渐降低, 这是笔者的新发现。在年时间尺度上, 对树干液流影响最大的为土壤温度和湿度, 能够解释94%的树干液流变化, 而与光照不存在显著相关关系。在日时间尺度上, 对树干液流影响最大的是土壤温度和光照, 其共同解释超过60%的树干液流变化(表 1)。在小时时间尺度上, 景观不同季节差异较大(表 3), 但综合分析结果发现:光照因子对对树干液流变化影响最大, 其次是空气温度和空气湿度(图 4)。
本文结果也表明:随着时间尺度的增大, 对树干液流影响逐渐由地上环境因子控制, 转换成为地下环境因子控制。比如, 在小时时间尺度上, 不同季节逐步回归分析结果中第1进入因子均为地上气象因子(光照和空气湿度), 第2进入因子5组中(春季、夏季、秋季、冬季和所有) 2组是地上气象因子(空气温度), 剩余为土壤生态因子(表 3)。而在日尺度上, 土壤温度首先进入, 其次是地上光照和空气温度(表 2)。月时间尺度上的结果, 存在显著相关性的2个因子均是地下生态因子(土壤温度和土壤湿度)(表 2)。这些结果可说明在较大时间尺度下, 与根系吸收水分直接相关的因子对树干液流影响更大, 即受水分供应相关过程限制, 而在较小的时间尺度下, 与叶片等耗散水分直接相关的生态因子对树干液流影响更大, 即受水分消耗相关过程限制。
当在不同时间尺度上对落叶松树干液流进行分析时, 所需要考虑的因素随时间尺度的增大而所需要考虑的因素减少, 这对于分析树干液流特征具有一定科学意义。在以月这个相对较大的时间尺度中, 1个环境因子(土壤温度)影响树干液流密度就能够解释树干液流变化的90%, 而在相对中等的时间尺度(天)条件下, 3个环境因子(土壤温度、光照和空气温度)显著影响树干液流, 解释其中62%的变化, 而在小时时间尺度下, 4个甚至全部5个因子均显著影响树干液流密度, 而且相关系数R2多在0.5以下。从分析中可以看出:在推算整株植物、甚至整个林分耗水量时, 在较大的尺度上进行较少量的环境因子监测, 即可得到精度更高的估计; 当需要对较小尺度上进行研究时, 依靠环境因子反推树干液流密度变化存在较大的风险, 最好直接使用TDP等先进仪器直接测定。
基于单株落叶松树干液流与环境因子相关关系的结果是否具有普遍性, 是值得怀疑的。为了进一步探讨上述结果是否具有普遍性, 对4株同时测定的落叶松树干液流数据进行了相关性分析。如果不同树木在相同生境下具有类似的树干液流特征, 不同个体间应该具有高度相关性, 笔者的结果也就应该具有普遍性。相关分析发现:不同个体之间的相关系数在0.88~0.96之间(表 5), 说明不同个体的树干液流密度年变化趋势趋于一致性。以单株落叶松的树干液流密度和环境因子的相关关系进行分析, 基本可以代表整个林分的其他落叶松个体。
5 结论不同时间尺度条件下, 影响树干液流的生态因子发生显著变化。在以月为时间尺度的结果中, 土壤温度和土壤湿度显著影响落叶松树干液流密度的变化, 能够解释超过93%的树干液流变化, 其他因子没有显著影响; 在以天为时间尺度中, 土壤温度为主要的影响因子, 解释树干液流变化的50%以上, 其次是光照和空气温度。在以小时为时间尺度中, 影响树干液流的因子较多, 而且不同季节差异性较大, 总体来讲, 地上因子包括光照和空气湿度的影响显著增强。在推算整株植物、甚至整个林分耗水量时, 在较大的尺度上进行较少量的环境因子监测, 即可得到精度更高的估计; 当需要对较小尺度上进行研究时, 依靠环境因子反推树干液流密度变化存在较大的风险, 最好直接使用TDP等先进仪器直接测定。
方精云. 2000. 全球变化生态学[M]. .北京: 高等教育出版社.
|
李海涛, 陈灵芝. 1998. 应用热脉冲技术对棘皮桦和五角枫树干液流的研究[J]. 北京林业大学学报, 20(1): 1-6. |
刘奉觉, EdwardsW R N, 郑世锴, 等. 1993. 杨树树干液流时空动态研究[J]. 林业科学研究, 6(4): 368-372. |
马玲, 赵平, 饶兴权, 等. 2005. 鹤山人工马占相思林水分生态研究[J]. 生态学报, 25(9): 2145-2151. |
孙迪, 关德新, 袁凤辉, 等. 2010. 辽西农林复合系统中杨树液流速率与气象因子的时滞效应[J]. 应用生态学报, 21(11): 2742-2748. |
孙鹏森, 马履一, 王小平, 等. 2000. 油松树干液流的时空变异性研究[J]. 北京林业大学学报, 22(5): 1-6. |
王华, 欧阳志云, 郑华, 等. 2009. 北京城区常见树种生长季树干液流的时滞特征[J]. 应用生态学报, 20(9): 2111-2117. |
王华田, 马履一. 2002. 利用热扩散式边材液流探针TDP测定树木整株蒸腾耗水量的研究[J]. 植物生态学报, 24(6): 661-667. |
王瑞辉, 奚如春, 徐军亮, 等. 2006. 用热扩散式茎流计测定园林树木蒸腾耗水量[J]. 中南林学院学报, 26(2): 7-12. |
王翠, 王传宽, 孙慧珍, 等. 2008. 移栽自不同纬度的兴安落叶松的树干液流特征[J]. 生态学报, 28(1): 136-144. |
谢恒星, 张振华, 杨润亚, 等. 2007. 龙爪槐树干液流相对于气象因子的滞后效应分析[J]. 林业科学, 43(5): 106-110. |
熊伟, 王彦辉, 徐德应. 2003. 宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应[J]. 林业科学, 39(2): 1-7. DOI:10.11707/j.1001-7488.20030201 |
尹光彩, 周国逸, 王旭, 等. 2003. 应用热脉冲系统对桉树人工林树液流通量的研究[J]. 生态学报, 23(10): 1984-1990. DOI:10.3321/j.issn:1000-0933.2003.10.005 |
尹光彩, 周国逸, MorrisJ, 等. 2004. 应用热脉冲系统对桉树人工林树液流通量的研究[J]. 浙江大学学报, 5(10): 1218-1225. DOI:10.3969/j.issn.1673-565X.2004.10.009 |
Anfodillo T, Sigalotti G B, Tomasi M, et al. 1993. Applications of a thermal imaging technique in the study of the ascent of sap in woody species[J]. Plant Cell and Environment, 16(8): 997-1001. DOI:10.1111/pce.1993.16.issue-8 |
Becker P. 1998. Limitations of a compensation heat pulse velocity system at low sap flow:implications for measurements at night and in shaded trees[J]. Tree Physiology, 18(3): 177-184. DOI:10.1093/treephys/18.3.177 |
Ford C R, Hubbard R M, Kloeppel B D, et al. 2007. A comparison of sap flux-based evapotranspiration estimates with catchment-scale water balance[J]. Agricultural and Forest Meteorology, 145(3/4): 176-185. |
Granier A. 1987. Evaluation of transpiration in a douglas fir stand by means of sap flow measurements[J]. Tree Physiology, 3(4): 309-320. DOI:10.1093/treephys/3.4.309 |
Granier A, Hucb R, Barigah S T. 1996. Transpiration of natural rain forest and its dependence on climatic factors[J]. Agricultural and Forest Meteorology, 78(1/2): 19-29. |
Granier A, Claustres J P. 1998. Water relations of a Norway spruce(Picea abies) tree growing in natural condition:Variation within the tree[J]. Acta Oecologica, 10(3): 295-310. |
Lindroth A, Cienciala E, Kucera J, et al. 1995. Sap flow by the heat balance method applied to small size Salix trees in a short rotation forest[J]. Biomass and Bioenergy, 8: 7-15. DOI:10.1016/0961-9534(94)00085-8 |
Nicolas E, Torrecillas A, Ortuno M F, et al. 2005. Evaluation of transpiration in adult apricot trees from sap flow measurements[J]. Agricultural Water Management, 72(2): 131-145. DOI:10.1016/j.agwat.2004.09.008 |
Swanson R H, Whitfield D W A. 1981. A number analysis of heat pulse velocity theory and practice[J]. Journal of Experimental Botany, 32(1): 211-239. |
Swanson R H. 1994. Significant historical developments in thermal methods for measuring sap flow in trees[J]. Agricultural and Forest Meteorology, 72(1/2): 113-132. |