 2012, Vol. 48
2012, Vol. 48文章信息
- 张立杰, 刘鹄
- Zhang Lijie, Liu Hu
- 祁连山林线区域青海云杉种群对气候变化的响应
- Response of Picea crassifolia Population to Climate Change at the Treeline Ecotones in Qilian Mountains
- 林业科学, 2012, 48(1): 18-21.
- Scientia Silvae Sinicae, 2012, 48(1): 18-21.
-
文章历史
- 收稿日期:2010-11-18
- 修回日期:2011-11-25
-
作者相关文章


2. 中国科学院寒区旱区环境与工程研究所临泽内陆河流域研究站 兰州 730000
2. Linze Inland River Basin Research station, Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences Lanzhou 730000
过去100年(1906—2005年),全球地表平均温度约升高0.74 ℃,20世纪80年代以来增温最明显(IPCC,2007)。气候变暖对全球自然生态系统产生了深远影响,作为全球陆地生态系统主体的森林对未来气候变化的响应更是人们关注的焦点。
高山林线对气候变化极为敏感,被认为是研究森林与气候变化关系最理想的场所。高山林线指示着森林分布的极限环境,多数研究认为热量或温度因子是控制林线形成的主要原因(王襄平等,2004)。许多研究证实,作为对气候变暖的回应,林线位置树木的高生长和年轮生长明显加快,林线种群密度的增加也清楚地显现出来,甚至在一些地区林线的位置也发生了明显的改变(Cuevas,2002; Klasner et al., 2002; Lloyd et al., 2003; 王晓春等,2004; 张桥英等,2007; Harsch,2009)。但由于受到树种和区域环境条件的影响,不同地区间林线种群对气候变化的响应程度存在很大差异(Holtmeier et al., 2005)。
许多研究已经证实祁连山在过去的50年里,平均气温出现显著变暖的趋势(贾文雄等,2008; 尹宪志等,2009)。青海云杉(Picea crassifolia)作为祁连山寒温性针叶林的唯一建群种,在维护区域生态安全方面发挥着重要作用。青海云杉林线种群是否也受到了区域气候变暖的影响,目前这方面的报道还比较少。本研究探讨祁连山青海云杉种群对区域气候变化的响应,并分析引起这些变化的主要原因,旨在为进一步开展半干旱山区植被对气候变化响应的研究提供必要的信息和数据积累。
1 研究区概况祁连山地处青藏、蒙新、黄土三大高原的交汇地带,属大陆性高寒半干旱、半湿润的森林草原气候。祁连山在过去的50年里,年平均气温以每10年0.30 ℃的倾向率增加,并且自20世纪80年代中期开始增温幅度明显加大(贾文雄等,2008; 尹宪志等,2009)。青海云杉是祁连山山地森林中分布最广、蓄积量最大的乔木树种,以不连续的斑块分布在海拔2 600~3 330 m的阴坡和半阴坡。植被随地形和气候的差异呈明显的垂直分布带,主要有山地森林草原带、亚高山灌丛草甸带及亚高山冰雪稀疏植被带。
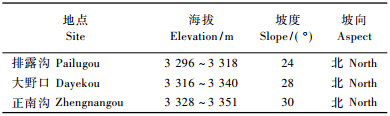
在祁连山中部的排露沟、大野口和正南沟阴坡林线位置,从青海云杉分布的树线位置,沿坡面向下各设置1块140 m×30 m的林线样地,样地的长边与坡向平行。各样地土壤和植被情况基本相似,青海云杉林下生长有以箭叶锦鸡儿(Caragana jubata)和吉拉柳(Salix gilashanica)等为主的高山灌丛,其盖度在0.6左右。样地土层厚40 ~60 cm,土壤以森林灰褐土为主。样地概况见表 1。
|
|
调查样地中所有青海云杉个体的相对位置、树高和胸径,对高度小于2.5 m的幼树,通过抽样调查轮生枝和芽鳞痕的方法确定其年龄,并建立树龄y和树高x(cm)的回归方程(y=0.111x+5.517,R2=0.928,P<0.01);对于枝轮无法确定的大树,抽样钻取树芯,建立树龄y和胸径x(cm)的回归方程(y=2.83x+32.18,R2=0.739,P<0.01),确定其树龄。并依据树龄重建近100年来林线样地内青海云杉的种群密度。
在每个样地内,对胸径大于10 cm的个体,在胸径部位各钻取1个树芯,共采集完整树芯82个,将树芯带回实验室经过风干、打磨,用树木年轮测定仪测定其年轮宽度值。采用ARSTAN年表研制程序,对样本序列的轮宽值进行去生长趋势(detrending)和标准化处理,建立标准化年表(STD)。利用Dendro2002对标准树木年轮指数与气候因子的关系进行相关分析(Biondi et al., 2004)。气象数据来源于祁连山区野牛沟气象站,气象站海拔3 180 m,距正南沟样地的直线距离为35 km。祁连山区气候垂直分带非常明显,选择海拔与样地接近的野牛沟气象站,可保证研究结果可靠性。在过去49年,野牛沟夏季和秋季平均气温均呈现显著上升趋势,尤其在过去29年(1979—2007),夏季和秋季升温趋势特别明显,年平均气温变化趋势也很明显,以每10年0.29 ℃的倾向率递增(图 1)。

|
图 1 野牛沟气象站近50年来夏季和秋季平均气温的变化 Figure 1 Variation of mean air temperature in summer and fall at Yeniugou weather station in past 50 years |
3块林线样地内,青海云杉林线种群均以幼龄个体为主(图 2),其中11~20龄的个体数量最多,占到总数的44.1%;1~30龄个体数量,达到了总数的80.9%,而60龄以上的个体数量较少。

|
图 2 青海云杉林线种群年龄结构 Figure 2 Age structure of Picea crassifolia population at treeline ecotones |
3块样地青海云杉林线种群密度在过去的90年里呈现明显的指数增长规律(图 3),进入1978年以后,种群密度开始大幅度增加,大约有80.9%的个体出现在近30年内。林线种群密度在过去的90年里,大约增长了23倍(由1918年的56株·hm-2增长到2008年的1 292株·hm-2)。

|
图 3 青海云杉林线种群密度的变化 Figure 3 Temporal change of Picea crassifolia population density at the treeline ecotones |
图 4根据林线样地内青海云杉个体的树龄和相对位置,重建了不同时期(50年前和100年前)林线样地内个体的相对位置,3个林线样地内青海云杉个体的时空分布特征基本相似,在靠近当前林线位置的区域,50年前和100年前仍然有部分青海云杉个体分布,这说明当前的林线位置在100年前已经形成,林线位置并没有发生实质性的位移。

|
图 4 不同时期青海云杉林线的相对位置 Figure 4 Relative position of Picea crassifolia trees at treeline ecotones in different periods x和y分别代表样地的边长; 较粗的黑色实线代表当前林线的位置。 x and y indicate the width and length of plots, the black bold line indicates the current timberline position of the plots. |
林线样地采集了82个树芯,建立了青海云杉树木年轮标准化年表(图 5),标准树轮宽度指数的平均敏感度为0.21,样本间的平均相关系数为0.69。树木年轮指数较好地反映了近238年来青海云杉径向生长变化的信息,在1770—1800年、1890—1930年和1950—1970年,青海云杉径向生长处于一个较低水平。20世纪30年代后期,其径向生长出现了一个较高水平的生长的过程,但持续时间不长。70年代后期至今,青海云杉径向生长出现了一个持续的高水平生长过程,这可能得益于其生长环境条件的持续改善。

|
图 5 青海云杉林线种群树木年轮指数的时间变化 Figure 5 Temporal change of tree ring width indices for Picea crassifolia population at treeline ecotones
—年轮指数的标准化年表序列Tree-ring width indices —年轮指数的10年滑动平均值10-year moving average of tree-ring width indices |
用Dendroclim 2002软件分析了林线青海云杉标准树轮宽度指数与单月(上年5月至当年10月)及各季(上年夏季至当年秋季)降水量和平均气温的相关性。青海云杉标准树轮宽度指数与当年夏季(5—7月)、秋季(8—10月)以及上一年夏季(5—7月)平均气温呈显著正相关(P<0.05),与上一年秋季及当年6月份的平均气温呈极显著正相关(P<0.01)。虽然与夏季各月降水量呈一定的正相关,但其相关性均未通过相关性检验。
4 结论与讨论青海云杉林线一般位于高海拔偏远山区的北坡,气候条件非常恶劣,祁连山青海云杉林线处根际土壤(10 cm深度)生长季平均温度仅为4.9 ℃,远低于全球林线生长季土壤平均温度(6.7 ℃)(胡启武等,2009)。在这种严酷的环境条件下,林线区域青海云杉对气候变暖具有更强的敏感性。祁连山在过去的49年里,气温出现显著变暖的趋势,与林线采样点最近的野牛沟气象站年平均气温以每10年0.29 ℃的倾向率递增,远高于全球每10年0.06 ℃的平均递增率(IPCC,2007)。温度的显著升高对林线植物光合作用具有正效应,有利于营养物质的积累,增加植物组织的木质化程度,进而增强林木对林线恶劣气候的抵御能力,提高林线植物的生长和幼苗的存活水平(Camarero et al., 1999; Batllori et al.,2008;王开运等,2010)。因此青海云杉林线种群密度增加与祁连山区域气候变暖密切相关。
祁连山大部分森林分布于高寒阴湿的半干旱山区,温度和水分条件是影响植物生长的主要限制性因子。祁连山区气候垂直分带非常明显,海拔每升高100 m,降水量增加18.6 mm,平均气温降低0.58 ℃(王金叶等,2001)。随海拔升高,水分条件会显著改善,而平均气温明显下降,因此在高海拔的林线区域,温度成为影响祁连山林线植物生长最主要的因子。已有研究表明:林线树木年轮一般会在夏季7—8月热量条件较好的50天内形成(Urbinati et al,1996)。青海云杉标准树轮宽度指数与当年夏季(5—7月)、秋季(8—10月)以及上一年夏季(5—7月)平均气温显著正相关,这可能与夏季和秋季平均气温升高可有效延长生长季有关。一些研究也证实,上一年夏季与秋季平均气温升高有利于光合产物积累,进而给来年植物的生长创造良好的物质条件(Takashashi et al., 2003; Wang et al., 2006)。
一般林线位置的波动大都是气候变化引起的,而这种波动只有在较大时间尺度上才能分辨出来(Klasner et al., 2002; MacDonald et al., 1998),而祁连山区气候出现明显变暖的趋势,也只是近30年来才发生的。如果祁连山区气候一直沿着持续变暖的趋势发展,青海云杉林线位置必将出现明显的上移。
胡启武, 吴琴, 张锋, 等. 2009. 祁连山青海云杉林树线温度特征[J]. 干旱区地理, 32(5): 698-703. |
贾文雄, 何元庆, 李宗省, 等. 2008. 祁连山区气候变化的区域差异特征及突变分析[J]. 地理学报, 63(3): 257-269. DOI:10.11821/xb200803004 |
王金叶, 常学向, 葛双兰, 等. 2001. 祁连山(北坡)水热状况与植被垂直分布[J]. 西北林学院学报, 16(Z1): 1-3. DOI:10.3969/j.issn.1001-7461.2001.z1.001 |
王开运, 胡庭兴, 张力, 等. 2010. 青藏高原东缘林线交错带糙皮桦幼苗光合特性对模拟增温的短期响应[J]. 植物生态学报, 34(3): 263-270. |
王襄平, 张玲, 方精云. 2004. 中国高山林线的分布高度与气候的关系[J]. 地理学报, 59(6): 871-879. DOI:10.11821/xb200406009 |
王晓春, 周晓峰, 李淑娟, 等. 2004. 气候变暖对老秃顶子林线结构特征的影响[J]. 生态学报, 24(11): 2412-2421. DOI:10.3321/j.issn:1000-0933.2004.11.010 |
尹宪志, 张强, 徐启运, 等. 2009. 近50年来祁连山区气候变化特征研究[J]. 高原气象, 28(1): 85-90. |
张桥英, 张运春, 罗鹏, 等. 2007. 白马雪山阳坡林线方枝柏种群的生态特征[J]. 植物生态学报, 31(5): 857-864. DOI:10.17521/cjpe.2007.0108 |
Batllori E, Gutiérrez E. 2008. Regional tree line dynamics in response to global change in the Pyrenees[J]. Journal of Ecology, 96(6): 1275-1288. DOI:10.1111/jec.2008.96.issue-6 |
Camarero J J, Gutierrez E. 1999. Structure and recent recruitment at alpine forest-pasture ecotones in the Spanish central Pyrenees[J]. écoscience, 6(3): 451-464. DOI:10.1080/11956860.1999.11682540 |
Cuevas J G. 2002. Episodic regeneration at the Nothofagus pumilio alpine timberline in Tierra del Fuego, Chile[J]. Journal of Ecology, 90(1): 52-60. DOI:10.1046/j.0022-0477.2001.00636.x |
Harsch M A, Hulme P E, McGlone M S, et al. 2009. Are treelines advancing? A global meta-analysis of treeline response to climate warming[J]. Ecology Letters, 12(10): 1040-1049. DOI:10.1111/ele.2009.12.issue-10 |
Holtmeier F K, Broll G. 2005. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales[J]. Global Ecology and Biogeography, 14(5): 395-410. DOI:10.1111/geb.2005.14.issue-5 |
IPCC. Climate change 2007: the physical science basis contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. New York: Cambridge University Press. https://link.springer.com/chapter/10.1007%2F978-1-4020-8709-7_20
|
Klasner F L, Fagre D B. 2002. A half century of change in alpine treeline patterns at Glacier National Park, Montana, USA[J]. Arctic, Antarctic, and Alpine Research, 34(1): 49-56. DOI:10.2307/1552508 |
Lloyd A H, Fastie C L. 2003. Recent changes in treeline forest distribution and structure in interior Alaska[J]. Ecoscience, 10(2): 176-185. DOI:10.1080/11956860.2003.11682765 |
MacDonald G M, Szeicz J M, Claricoates J, et al. 1998. Response of the central Canadian treeline to recent climatic changes[J]. Annals of the Association of American Geographers, 88(2): 183-208. DOI:10.1111/1467-8306.00090 |
Takahashi K, Azuma H, Yasue K. 2003. Effects of climate on the radial growth of tree species in the upper and lower distribution limits of an altitudinal ecotone on Mount Norikura[J]. Ecol Res, 18(5): 549-558. DOI:10.1046/j.1440-1703.2003.00577.x |
Urbinati C, Carrer M, Anfodillo T. 1996. Dendroecology at thetimberline: growth dynamics and climatic factors[J]. Dendronatura, 17(1): 41-50. |
Wang T, Zhang Q, Ma K. 2006. Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China[J]. Global Ecol Biogeogr, 15(4): 406-415. DOI:10.1111/geb.2006.15.issue-4 |