 2012, Vol. 48
2012, Vol. 48文章信息
- 任学敏, 杨改河, 秦晓威, 王得祥, 刘振学, 赵双喜, 白宇
- Re Xuemin, Yang Gaihe, Qin Xiaowei, Wang Dexiang, Liu Zhenxue, Zhao Shuangxi, Bai Yu
- 巴山冷杉-牛皮桦混交林乔木更新及
- Tree Regeneration in an Abies fargesii-Betula utilis Mixed Forest and Effects of the Soil Chemical Properties on Regeneration
- 林业科学, 2012, 48(1): 1-6.
- Scientia Silvae Sinicae, 2012, 48(1): 1-6.
-
文章历史
- 收稿日期:2010-08-31
- 修回日期:2011-11-25
-
作者相关文章



2. 陕西省循环农业工程技术研究中心 杨凌 712100;
3. 西北农林科技大学林学院 杨凌 712100
2. Research Center of Recycle Agricultural Engineering and Technology of Shaanxi Province Yangling 712100;
3. College of Forestry, Northwest A & F University Yangling 712100
森林植物更新尤其是乔木更新对种群的增殖、扩散和延续及群落发展具有重要作用(Herrera,1995; 李小双等,2007)。乔木更新可细分为林下更新和林隙更新,二者均受到各种环境条件的影响,土壤化学性质就是重要因素之一。Catovsky等(2002)研究发现幼苗的存活和生长与土壤pH值有关; Bungard等(2002)认为,土壤氮素含量是幼苗生长的限制因子之一; 闫海冰等(2010)研究表明,土壤氮素含量高的区域,云杉(Picea asperata)更新苗数量多; Muscolo等(2007)指出,土壤C,N和P含量对乔木更新有重要影响。因此,了解土壤化学性质对乔木更新的影响对于了解森林动态具有重要意义。
太白山巴山冷杉(Abies fargesii)-牛皮桦(Betulautilis)混交林分布于海拔2 450~3 050 m,该海拔范围还分布有大面积的巴山冷杉纯林、红桦(Betula albosinensis)纯林和牛皮桦纯林(刘慎谔,1985)。长期以来,学者们对这一植被带进行了较多研究(朱志诚,1991; 傅志军等,1994a; 1994b;岳明等,2000; 2002;林玥等,2009),但主要集中于红桦纯林、牛皮桦纯林和巴山冷杉纯林,而对该区域大面积分布的巴山冷杉-牛皮桦混交林的研究则少有报道,对巴山冷杉-牛皮桦混交林乔木更新及土壤化学性质对更新影响的研究未见报道。本研究调查巴山冷杉-牛皮桦混交林林下和林隙的乔木更新状况,并探索土壤化学性质及林隙大小对乔木更新的影响,以期为理解该区森林动态提供理论依据,为当地森林经营管理提供理论指导。
1 研究区概况研究于秦岭太白山自然保护区(107°41'23″—107°51'40″E,33°49'31″—34°08'11″N)南坡的巴山冷杉-牛皮桦混交林内进行。该区受东南季风影响,雨量较充沛,厚畛子气象站(距样地约15 km)25年的观测数据显示,该区年均降水量945.5 mm,年均气温8.4 ℃,最冷月1月均温-4.2 ℃,最热月7月均温20.4 ℃。极端低温-19 ℃,极端高温29.7 ℃。土壤类型为山地暗棕壤,成土母岩主要是片麻岩和花岗岩(雷梅等,2001)。林下灌木层植物主要有四川忍冬(Lonicera szechuanica)、菰帽悬钩子(Rubus pileatus)、香柏(Sabina squamata var.wilsonii)和蒙古绣线菊(Spiraea mongolica); 草本层植物主要有大毛状薹草(Carex capilliformis var.major)、太白山槖吾(Ligulara dolichobotrys)、裸茎碎米荠(Cardamine scaposa)、山酢浆草(Oxalisacetosella subsp. griffithii)、大耳叶风毛菊(Saussureamacrota)和翅茎风毛菊(Saussurea cauloptera)。
2 研究方法调查及取样时间为2009年7—9月和2010年7—8月。在巴山冷杉-牛皮桦混交林内建立3条样带,间隔为50 m,样带长500 m、宽20 m。在每条样带上每隔30 m设1块20 m×20 m样地(取样地时尽量避开较大林隙),3条样带共设30块样地。用手持式GPS500测定每块样地的海拔,用地质罗盘测量坡度和坡向,目测植被盖度,记录每块样地中出现的乔木物种名,估计每个物种的盖度,记录每块样地乔木(幼苗: 0.1 m≤H<1.3 m; 幼树: 1.3 m ≤ H<7.5 m,DBH<7.5 cm; 成年树:H≥7.5 m,DBH≥7.5 cm。H为株高)的株数并估算每株高度,用胸径尺测量所有株高大于1.3 m乔木的胸径。
在每条样带内或其附近(距样带20 m范围内)分别随机调查10个林隙,均为扩展林隙,其定义同Runkle(1982),共有30个林隙被调查。林隙边界由胸径大于7.5 cm、高度大于7.5 m的边界木来确定。测量距离最远的2株边界木的距离(L)和垂直于最大长度的最近的2株边界木的距离(W),用计算椭圆面积(A)公式(A=πLW/4) 近似计算林隙面积(Nagel et al., 2010)。用前述方法记录每个林隙中出现的乔木种名、株数、株高(≥0.1 m)和胸径。
收集每块样地和每个林隙0~30 cm土层的土壤样品。在样地四角及中心各挖1个剖面(土壤厚度不足30 cm的以挖到岩石为止,并测量土壤厚度,以5个剖面土壤厚度均值作为该样地土壤厚度),沿剖面从上到下收集土壤,将每块样地收集的土壤混合为1个土样; 视林隙大小在每个林隙挖3~5个剖面(土壤厚度不足30 cm的以挖到岩石为止,并测量土壤厚度,以林隙内各剖面土壤厚度均值作为该林隙土壤厚度),沿剖面从上到下收集土壤,将每个林隙收集的土壤混合为1个土样。样地和林隙各收集了30个土壤样品。将土壤样品带回实验室,pH值用pHS-3CT型pH计测定,有机质含量用重铬酸钾容量法-外加热法测定,全N含量用凯氏定氮法测定,碱解N含量用碱解扩散法测定,全P含量用酸溶-钼锑抗比色法测定,有效P含量用NH4 FHCl浸提法测定。
两尾t测验用于检验林下、林隙内更新苗密度及土壤化学性质的差异显著性; 皮尔逊相关用于分析林下、林隙内更新苗密度与其相应土壤化学性质的关系、林隙大小与更新苗密度间的关系。图由Origin 7.5完成,数据统计分析由SPSS 17.0完成
3 结果与分析 3.1 样地特征从表 1可以看出:样地海拔、坡度、坡向、土壤厚度和植被盖度的标准差均较小,表明研究区的环境特征较为一致。调查中仅发现2个乔木物种,即巴山冷杉和牛皮桦,巴山冷杉在研究区域占主导地位。样地乔木组成特征见表 2。
|
|

|
|

无论林下还是林隙内,巴山冷杉的幼苗和幼树都占绝对优势(图 1)。整体上,巴山冷杉-牛皮桦混交林更新密度不大,为0.059株·m-2,其中巴山冷杉更新密度为0.046株·m-2,占78.1%,牛皮桦为0.013株·m-2,占28.9%。林下与林隙内更新密度差异不明显,巴山冷杉在林下和林隙内的更新密度分别为0.045和0.052株·m-2,牛皮桦更小,分别为0.013和0.012株·m-2。在林下,巴山冷杉是幼苗少、幼树多,而在林隙内情况恰恰相反; 牛皮桦在林下与林隙内均是幼苗多、幼树少(图 1)。虽然巴山冷杉、牛皮桦总更新苗密度在林下、林隙内差异不显著(巴山冷杉,t=-1.543,P=0.128;牛皮桦,t=-0.119,P=0.906),但巴山冷杉在林下与林隙内的幼苗密度差异显著(t=-2.630,P=0.012),其余不显著(巴山冷杉幼树t=0.476,P=0.636;牛皮桦幼苗t=-0.499,P=0.619;牛皮桦幼树t=1.292,P=0.202)。

|
图 1 林下、林隙内乔木更新苗密度 Figure 1 Densities of tree regeneration in understory and gap |
相关分析表明:总的更新苗(巴山冷杉+牛皮桦)、幼苗(巴山冷杉+牛皮桦)、幼树(巴山冷杉+牛皮桦)密度与林隙面积存在负相关关系,但均不显著(分别为: r=-0.342,P=0.065;r=-0.210,P=0.265;r=-0.338,P=0.068)(图 2); 巴山冷杉幼树及更新苗密度与林隙面积呈显著负相关(分别为: r=-0.399,P=0.029;r=-0.397,P=0.030),巴山冷杉幼苗、牛皮桦幼树、牛皮桦幼苗及牛皮桦更新苗密度与林隙面积的相关性均不显著。

|
图 2 林隙面积与更新苗密度的相关性 Figure 2 Correlation between gap size and regeneration densities |
在林下,巴山冷杉幼苗密度与土壤pH值、有机质含量、碱解N含量、全P含量和有效P含量负相关,但均未达到显著水平; 巴山冷杉幼树密度与土壤pH值、有机质含量、全N含量、碱解N含量、全P含量和有效P含量负相关,但均未达到显著水平; 牛皮桦幼苗密度和幼树密度均与土壤pH值、有机质含量、全N含量、碱解N含量、全P含量和有效P含量正相关,其中幼树密度与土壤pH值和全P含量的相关达极显著水平,相关系数分别为0.599和0.517,幼树密度与碱解N含量、幼苗密度与土壤pH值和全P含量的相关性达显著水平,相关系数分别为0.362,0.398和0.418(表 3)。
|
|
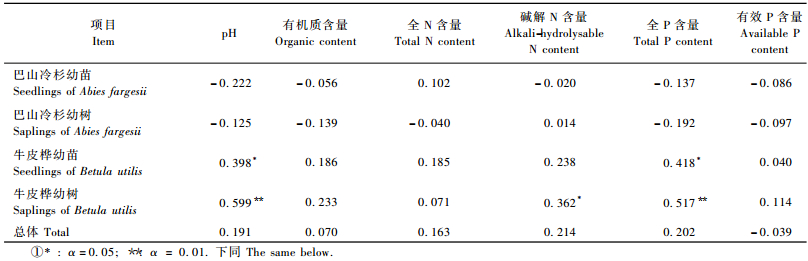
在林隙内,乔木更新苗密度与土壤全P和有效P含量呈显著负相关,相关系数分别为-0.438和-0.390。巴山冷杉幼苗密度与土壤全P含量、幼树密度与土壤有效P含量均呈显著负相关,相关系数分别为-0.392和-0.364,巴山冷杉幼树密度和牛皮桦幼树密度与土壤全P含量极显著负相关,相关系数分别为-0.486和-0.592(表 4)。
|
|
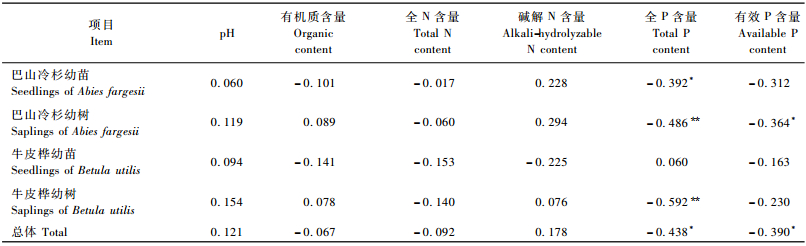
尽管林下、林隙内更新苗密度与土壤各化学指标之间的相关性多数没达到显著水平,但依然能从这些相关关系中看出林下巴山冷杉更新苗密度与土壤各化学指标基本呈负相关,而牛皮桦更新苗密度与之呈正相关(表 3)。整体看来,林隙内的相关关系比林下要复杂(表 4),这在一定程度上反映了林隙小环境比林下更为异质的特点。
3.5 林下、林隙内土壤化学性质对比林下土壤pH值高于林隙内,且达到显著水平,而有机质、全N、碱解N、全P和有效P含量均低于林隙,且差异均达到了显著水平(表 5)。
|
|

众多研究表明:林隙的形成创造了不同于林下的小生境,从而对树木更新产生重要影响(陈志刚等,2005; Dai,1996; Duan et al., 2009; Abd Latif et al.,2010)。Fraver等(2008)研究认为,林隙更利于树木更新; Denslow(1995)和Busing等(1997)的研究也证实了林隙内有比林下更高的树木更新密度。本研究也发现,林隙内乔木更新密度高于林下,但差异并没有达到显著水平,这说明本研究区林隙并没有强烈影响乔木更新。调查发现,巴山冷杉和牛皮桦更新苗数量均较少,尤其是牛皮桦,其更新苗密度仅为0.013株·m-2,反映了其天然更新不良,这与前人的研究结果相一致(李家俊等,1989; 傅志军等,1994b)。
虽然林隙内更新苗密度与林下整体上差异不显著,但林隙内巴山冷杉更新苗密度显著高于林下(图 1),而又与林隙面积之间呈显著负相关(图 2),说明较小林隙更利于巴山冷杉更新,这与前人的研究结果近似(Huth et al., 2006; 符婵娟等,2009)。小林隙更有利于巴山冷杉更新,其可能原因是小林隙周围高大的边界木使林隙内依然处于相对较阴暗环境,与林下相比又由于竞争的减少(McCarthy,2001),更多资源能用于更新,这些均为耐荫的巴山冷杉更新提供了有利条件(巴山冷杉更新苗平均密度在小林隙(<50 m2)为0.096株·m-2); 而中(50~100 m2)、大(>100 m2)林隙虽然也有充足的资源,但光照明显强于小林隙(Gálhidy et al., 2006; Abd Latif et al., 2010),这可能会抑制巴山冷杉更新。牛皮桦更新苗密度在林隙内与林下差异不显著,并且与林隙大小也没有显著相关关系,较大林隙内其更新苗密度还有下降趋势(在小、中、大林隙内的密度分别为0.010,0.020和0.008株·m-2),说明林隙不是牛皮桦更新的制约因素。
土壤是植物更新的基质,其化学性质是影响植物更新的关键因素之一。Stevenson等(1986)、Muscolo等(2007)指出,土壤酸碱度的大小及营养元素的缺乏可对植物的更新、生长产生直接影响。在林下内,土壤pH值与牛皮桦更新苗密度存在显著正相关(表 3),说明牛皮桦更适于低酸的环境; 碱解N和全P含量也与牛皮桦的更新存在显著正相关(表 3),说明碱解N和全P含量的不足可能对牛皮桦的更新产生一定的抑制。本研究发现,在林隙,影响更新的主要是全P和有效P含量,其中土壤全P含量与巴山冷杉幼树密度和牛皮桦幼树密度均呈极显著负相关,全P含量与巴山冷杉幼苗密度、有效P含量与牛皮桦幼树密度呈显著负相关(表 4)。这里尤其值得注意的是,土壤全P含量对牛皮桦幼树密度的影响由林下的显著正相关变为了林隙的显著负相关(表 3,4)。那么,适宜牛皮桦更新的土壤全P含量可能为(184.22±35.79)~(322.28±86.13) mg·kg-1(表 5); 林隙土壤全P和有效P含量可能对巴山冷杉的更新产生了一定抑制,但适宜巴山冷杉更新的P的浓度范围是多少,还需进一步研究。
陈志刚, 樊大勇, 张旺锋, 等. 2005. 林隙与林下环境对锐齿槲栎和米心水青冈种群更新的影响[J]. 植物生态学报, 29(3): 354-360. DOI:10.17521/cjpe.2005.0046 |
符婵娟, 刘艳红, 赵本元. 2009. 神农架巴山冷杉群落更新特点及影响因素[J]. 生态学报, 29(8): 4179-4186. |
傅志军, 郭俊理. 1994a. 太白山红桦林的初步研究[J]. 植物生态学报, 18(3): 261-270. |
傅志军, 郭俊理, 孙宗明. 1994b. 秦岭太白山牛皮桦林初步研究[J]. 宝鸡文理学院学报:自然科学版, (1): 79-83. |
雷梅, 常庆瑞, 冯立孝, 等. 2001. 太白山土壤特性及氧化铁发生学特征[J]. 地理研究, 20(1): 83-90. |
李家俊, 赵一庆, 薄颖生, 等. 1989. 太白山自然保护区主要森林植被类型调查∥李家俊. 太白山自然保护区综合考察论文集. 西安: 陕西师范大学出版社, 141-158.
|
李小双, 彭明春, 党承林. 2007. 植物自然更新研究进展[J]. 生态学杂志, 26(12): 2081-2088. |
林玥, 任坚毅, 岳明. 2009. 太白山巴山冷杉-牛皮桦混交林空间格局及关联性研究[J]. 武汉植物学研究, 27(1): 47-54. |
刘慎谔. 1985. 刘慎谔文集[M]. 北京: 科学出版社: 74-85.
|
闫海冰, 韩有志, 杨秀清, 等. 2010. 关帝山云杉天然更新与土壤有效氮素异质性的空间关联性[J]. 应用生态学报, 21(3): 533-540. |
岳明, 任毅, 党高弟, 等. 2000. 佛坪国家级自然保护区植被垂直带谱及其与邻近地区的比较[J]. 武汉植物学研究, 18(5): 375-382. |
岳明, 张林静, 党高弟, 等. 2002. 佛坪国家级自然保护区植物群落物种多样性与海拔梯度的关系[J]. 地理科学, 22(3): 349-354. |
朱志诚. 1991. 秦岭太白山桦林的稳定性[J]. 武汉植物学研究, 9(2): 169-175. |
Abd Latif Z, Blackburn G A. 2010. The effects of gap size on some microclimate variables during late summer and autumn in a temperate broadleaved deciduous forest[J]. International Journal of Biometeorology, 54(2): 119-129. DOI:10.1007/s00484-009-0260-1 |
Bungard R A, Zipperlen S A, Press M C, et al. 2002. The influence of nutrients on growth and photosynthesis of seedlings of two rainforest dipterocarp species[J]. Functional Plant Biology, 29(4): 505-515. DOI:10.1071/PP01137 |
Busing R T, White P S. 1997. Species diversity and small-scale disturbance in an old-growth temperate forest: a consideration of gap partitioning concepts[J]. Oikos, 78(3): 562-568. DOI:10.2307/3545618 |
Catovsky S, Bazzaz F A. 2002. Feedbacks between canopy composition and seedling regeneration in mixed conifer broad-leaved forests[J]. Oikos, 98(3): 403-420. DOI:10.1034/j.1600-0706.2002.980305.x |
Dai X B. 1996. Influence of light conditions in canopy gaps on forest regeneration: a new gap light index and it's application in a boreal forest in east-central Sweden[J]. Forest Ecology and Management, 84(1/3): 187-197. |
Denslow J S. 1995. Disturbance and diversity in tropical rainforests: the density effect[J]. Ecological Applications, 5(4): 962-968. DOI:10.2307/2269347 |
Duan W B, Wang J, Li Y. 2009. Microenvironmental heterogeneity of physical soil properties in a broad-leaved Pinus koraiensis forest gap[J]. Frontiers of Forestry in China, 4(1): 38-45. DOI:10.1007/s11461-009-0018-2 |
Fraver S, Jonsson B G, Jönsson M, et al. 2008. Demographics and disturbance history of a boreal old-growth Picea abies forest[J]. Journal of Vegetation Science, 19(6): 789-798. DOI:10.3170/2008-8-18449 |
Gálhidy L, Mihók B, Hagyó A, et al. 2006. Effects of gap size and associatedchanges in light and soil moisture on the understorey vegetation of Hungarian beech forest[J]. Plant Ecology, 183(1): 133-145. DOI:10.1007/s11258-005-9012-4 |
Herrera J. 1995. Acorn predation and seedling production in a low density population of cork oak(Quercus suber L.)[J]. Forest Ecology and Management, 76(1/3): 197-201. |
Huth F, Wagner S. 2006. Gap structure and establishment of Silver birch regeneration(Betula pendula Roth.)in Norway spruce stands(Picea abies L. Karst.)[J]. Forest Ecology and Management, 229(1/3): 314-324. |
McCarthy J. 2001. Gap dynamics of forest trees: a review with particular attention to boreal forests[J]. Environmental Reviews, 9(1): 1-59. DOI:10.1139/a00-012 |
Muscolo A, Sidari M, Mercurio R. 2007. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine(Pinus laricio, Poiret)stands[J]. Forest Ecology and Management, 242(2/3): 412-418. |
Nagel T A, Svoboda M, Rugani T, et al. 2010. Gap regeneration and replacement patterns in an old-growth Fagus-Abies forest of BosniaHerzegovina[J]. Plant Ecology, 208(2): 307-318. DOI:10.1007/s11258-009-9707-z |
Runkle J R. 1982. Patterns of disturbance in some old-growth mesic forests of eastern North America[J]. Ecology, 63(5): 1533-1546. DOI:10.2307/1938878 |
Stevenson J J, Cole M A. 1986. Cycles of soil: carbon, nitrogen, phosphorus, sulfur, micronutrients[M]. New York: John Wiley & Sons: 279-325.
|