 2011, Vol. 47
2011, Vol. 47文章信息
- 阎伟, 宗世祥, 王荣, 王建伟, 曹川建, 骆有庆
- Yan Wei, Zong Shixiang, Wang Rong, Wang Jianwei, Cao Chuanjian, Luo Youqing
- 油蒿不同演替阶段钻蛀性害虫幼虫与天敌在空间格局上的关系
- Spatial Pattern Relationships Between Woodborer Larvae and Their Natural Enemies in Artemisia ordosica Shrubs
- 林业科学, 2011, 47(12): 179-183.
- Scientia Silvae Sinicae, 2011, 47(12): 179-183.
-
文章历史
- 收稿日期:2009-12-02
- 修回日期:2010-09-04
-
作者相关文章



2. 中国热带农业科学院椰子研究所 文昌 571339;
3. 宁夏森林病虫害防治检疫总站 银川 750004
2. Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences Wenchang 571339;
3. Forest Diseases and Insect Pests Control and Quarantine Station of Ningxia Yinchuan 750004
油蒿(Artemisia ordosica)钻蛀性害虫包括沙蒿圆吉丁虫(Sphenoptera sp.)(鞘翅目:吉丁虫科)、沙蒿大粒象(Adosopius sp.)(鞘翅目:象甲科)、沙蒿木蠹蛾(Holcocerus artemisiae)(鳞翅目:木蠹蛾科)和一种卷蛾科(Tortricidae)蛀茎害虫,主要分布于我国的内蒙古、陕西、宁夏、甘肃等省区(阎伟等,2009a; 2009b),且有大面积暴发的趋势。它们主要以幼虫危害油蒿根和茎基部,在根茎内钻蛀隧道,破坏木质部而导致油蒿死亡。因其只在成虫期行裸露性生活,营隐蔽性生活时间很长,监测和防治都极为困难。油蒿是我国特有的优良固沙半灌木植物,亦称黑沙蒿,属菊科植物,是毛乌素沙地中分布最广泛的沙生植物群落的优势种,遍见于半固定与固定沙丘沙地,是沙地天然植物群落,具有耐沙埋、抗风蚀、耐贫瘠、分枝和结实性能好的特性,对毛乌素沙地系统的稳定起着关键作用(北京大学地理系等,1983; 张新时,1994; 杨洪晓等,2004; 张德魁等,2007)。目前对油蒿钻蛀性害虫的生物学和生态学方面的研究报道不多。宁夏盐池县油蒿钻蛀性害虫的主要天敌为寄生于幼虫的一种蒲螨(Pyemotes herfsi)(蒲螨科Pyemotidae)(马立芹等,2008)和啄开根部捕食幼虫及蛹的斑翅山鹑(Perdix dauuricae)。为探讨合理有效的综合治理技术,2006—2008年在宁夏盐池县对4种油蒿钻蛀性害虫及其寄生性天敌蒲螨和斑翅山鹑啄痕的空间格局进行系统调查。
1 研究地概况与方法 1.1 研究地概况调查地选在近年来受油蒿钻蛀性害虫危害比较严重的宁夏中东部的盐池县。该县位于毛乌素沙地西南缘,属于温带半干旱季风气候,年均温6.0~8.5 ℃,年均降水量250~350 mm,且主要集中在7—9月(占年降水量的68.3%),年均蒸发量2 000 mm以上。地表形态主要表现为梁地与滩地、沙丘与滩地相间分布。土壤主要有沙地上的各类风沙土,以及梁地上的栗钙土或淡栗钙土。调查地基本情况:半固定沙丘,总盖度≤ 20%,油蒿盖度13.9%,地表几乎无结皮; 植被主要以油蒿为主,仅有沙米(Agirophyllum pungens)、虫实(Corispermum heptapotamicum)等少量伴生种。固定沙丘,总盖度25%~50%,油蒿盖度26.1%,地表具灰白色降尘结皮并逐渐演变为土壤生物结皮; 植被以油蒿为建群种,伴生种主要有糙隐子草(Cleistogenes squarrosa)、兴安胡枝子(Lespedeza davurica)、草木樨状黄芪(Astragalus melilotoides)、乳浆大戟(Euphorbia esula)、雾冰藜(Bassia dasyphylla)、沙葱(Allium prostratum)、赖草(Leymus secalinus)等。生草沙丘,总盖度35%~60%,油蒿盖度31.4%,地表具发育良好的土壤生物结皮; 植被以油蒿为建群种,常见种主要为狗尾草(Setaria viridis)、画眉草(Eragrostis pilosa)、刺藜(Chenopodium aristatum)、山苦荬菜(Ixeris chinensis)、乳浆大戟、地锦(Euphorbia humifusa)等杂草(北京大学地理系等,1983; 王妍等,2006)。
1.2 调查方法在林分踏查的基础上,根据油蒿群落的演替特点(北京大学地理系等,1983; 王庆锁等,1997; 郭柯,2000; 张军等,2007),选择3种不同类型的典型油蒿群落为试验地,即半固定沙丘试验地、固定沙丘试验地和生草沙丘试验地,依次记为Ⅰ、Ⅱ、Ⅲ区,合计面积约6 km2。在每个区中,根据钻蛀性害虫对油蒿的危害程度,分别又选取了16, 18和27块样方,每个样方的大小为10 m×10 m(CCICCD,1997; 何志斌等,2004),这些样方均匀分布在3个调查区内。
采用双对角线抽样方法,从每个样方内抽取20丛油蒿,进行全株解剖。详细记录每丛油蒿的长势、冠幅、主根粗、树龄、是否受害、钻蛀性害虫的虫口数量、被寄生数和斑翅山鹑啄痕数量及寄生部位等指标。往年死树和旧痕不计入。
1.3 空间分布参数参照丁岩钦(1994)和邬祥光(1985)介绍的部分指标判别昆虫在油蒿各个演替阶段的空间分布情况。
1) 扩散系数C=S2/x。其中,S2表示样本方差,x表示样本平均值。C<1为均匀分布; C=1为随机分布; C>1为聚集分布。
2) 平均拥挤度m*=x+I及聚块性指标m*/x。m*、m*/ x<1为均匀分布; m*、m*/x=1为随机分布; m*、m*/x>1为聚集分布。
3) 负二项分布k=x2/(S2-x)。k值越小,聚集度越大,k→0时高度聚集; k值越大,聚集度越小,k→∞时(一般>8)则逼近随机分布。
4) 种群聚集均数λ=(x/2k)/γ。式中γ为自由度为2k时的X0.052值。λ<2表示聚集由环境因子引起的; λ≥2表示聚集由本身习性引起的。
5) 个体群大小指标 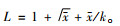
据
采用SPSS11.5软件对所有试验数据进行统计分析(余建英等,2003)。
2 结果与分析 2.1 水平方向上的空间分布规律1) 油蒿钻蛀性害虫幼虫(表 1) 沙蒿圆吉丁虫、沙蒿大粒象、沙蒿木蠹蛾和一种卷蛾科蛀茎害虫幼虫的扩散系数C、平均拥挤度m*、聚块性指标m*/X均大于1,负二项分布k值较小,在油蒿群落演替经历的3个阶段中,4种油蒿钻蛀性害虫幼虫在水平方向上均呈聚集型分布,但负二项分布k值在演替过程中均表现为逐渐增大,表明随着油蒿群落的演替,以上4种钻蛀性害虫的聚集度有减弱的趋势,其中尤其油蒿演替的生草阶段最为明显; 此外,随着4种油蒿钻蛀性害虫虫口密度的变化,其各自聚集的个体群(L)也相应发生变化; 种群聚集均数λ均大于2,说明4种油蒿钻蛀性害虫幼虫聚集分布的原因是由其自身习性引起的。
|
|
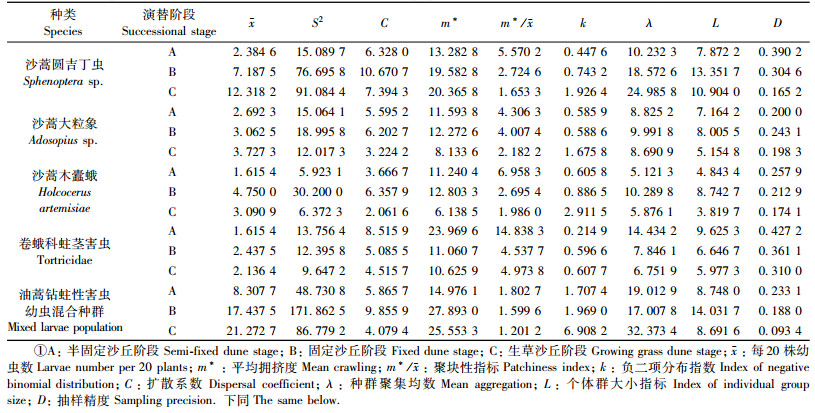
从表 1还可以看出:油蒿钻蛀性害虫混合种群幼虫在每个演替阶段的取样精度均符合要求(D<0.25),扩散系数C、平均拥挤度m*、聚块性指标m*/x均大于1,说明油蒿钻蛀性害虫幼虫在每个演替阶段的水平分布均为聚集型分布,种群聚集均数λ均大于2,说明油蒿钻蛀性害虫幼虫聚集分布的原因是由其自身习性引起的。
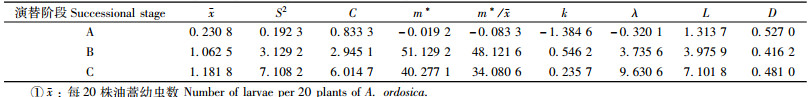
2) 被蒲螨寄生的害虫幼虫(表 2) 被蒲螨寄生的钻蛀性害虫幼虫的水平分布与其相应的株平均虫口密度和油蒿演替阶段有关,当每20株油蒿害虫平均密度在0.25头以下时为均匀分布,而在0.25头以上时则为聚集分布; 一般来说,在油蒿演替的半固定沙丘阶段,平均密度在0.25头以下,而在固定沙丘和生草沙丘阶段,该值都在1头以上。
|
|
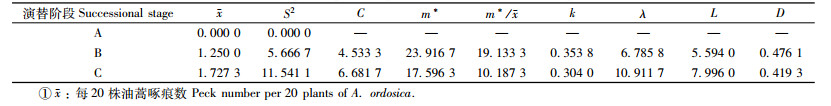
3) 斑翅山鹑 斑翅山鹑一年四季都对当地的油蒿钻蛀性害虫发挥控制作用(表 3)。在半固定沙丘阶段没有发现啄痕,在固定沙丘和生草沙丘阶段,各样地斑翅山鹑啄食形成的啄痕均呈聚集型分布,这显然与其在油蒿林中的主要捕食猎物油蒿钻蛀性害虫的分布模式密切相关。
|
|
被油蒿钻蛀性害虫危害的油蒿不同高度虫口数量频数分布的分析表明(图 1):4种油蒿钻蛀性害虫在油蒿演替的各个阶段内,从地上部6 cm至地下12 cm的范围内均有分布,主要集中于地上3 cm至地下6 cm的根茎部,大至呈抛物线趋势型; 在生草沙丘阶段4种油蒿钻蛀性害虫的数量最多,半固定沙丘阶段最少。虽然在油蒿的演替过程中,油蒿群落的物种组成、多样性、丰富度及周围的环境都会发生一系列的变化(张新时,1994),但由于钻蛀性害虫隐蔽生活的习性,上述4种油蒿钻蛀性害虫在油蒿演替的不同阶段,在油蒿垂直高度上的分布没有差异(P>0.05)。

|
图 1 沙蒿圆吉丁虫(a)、沙蒿大粒象(b)、沙蒿木蠹蛾(c)和卷蛾科蛀茎害虫(d)幼虫的垂直分布 Figure 1 Vertical distribution of larvae of Sphenoptera sp.(a), Adosopius sp.(b), Holcocerus artemisiae (c) and Tortricidae(d) A:半固定沙丘阶段Semi-fixed dune stage; B:固定沙丘阶段Fixed dune stage; C:生草沙丘阶段Growing grass dune stage. |
其中,沙蒿圆吉丁虫主要分布在地表至地下3 cm的根部,占沙蒿圆吉丁虫幼虫总数的66.3%;沙蒿大粒象主要分布于地下3 cm至地下6 cm的根部,占沙蒿大粒象幼虫总数的53.4%;沙蒿木蠹蛾主要分布于地表至地下6 cm的根部,占沙蒿木蠹蛾幼虫总数的83.2%;卷蛾科害虫则主要分布在地表至地下3 cm的根部,占卷蛾科害虫幼虫总数的66.7%。此外,通过野外调查发现在树龄较大、根部较粗的油蒿上,沙蒿大粒象和沙蒿木蠹蛾可以分布于地下12 cm以下的根部。
被蒲螨寄生的油蒿钻蛀性害虫幼虫主要分布在生草沙丘阶段,其垂直分布规律与上述4种油蒿钻蛀性害虫的垂直分布规律相似,大至呈抛物线趋势型。其中,79.3%的被蒲螨寄生的油蒿钻蛀性害虫幼虫分布在地表至地下3 cm的根部(图 2),说明蒲螨在三维空间上对油蒿钻蛀性害虫有一定的追随作用,籍此提高蒲螨对油蒿钻蛀性害虫的寄生率。

|
图 2 被蒲螨寄生的幼虫在油蒿干基和根部的垂直分布 Figure 2 Vertical distribution of parasitized larvae of woodborer by P. herfsi in stem basal and root of A. ordosica |
对油蒿钻蛀性害虫分布与天敌分布关系的分析表明:在油蒿群落由半固定沙丘阶段向生草沙丘阶段演替的过程中,油蒿钻蛀性害虫的数量逐渐增加; 在此过程中,被蒲螨寄生的油蒿钻蛀性害虫的数量也逐渐增多,但是在固定沙丘阶段和生草沙丘阶段被蒲螨寄生的油蒿钻蛀性害虫差异不显著(P>0.05);在半固定沙丘阶段,没有发现斑翅山鹑的啄痕,在固定沙丘阶段和生草沙丘阶段均发现斑翅山鹑的啄痕,而且固定沙丘阶段的啄痕数显著多于生草沙丘阶段的啄痕数(P<0.05)(表 4)。
|
|

本研究分别对经历半固定沙丘、固定沙丘和生草沙丘演替阶段的油蒿进行全株解剖,研究不同演替阶段油蒿钻蛀性害虫幼虫的三维分布。结果表明:在油蒿群落演替的每个阶段,4种钻蛀性害虫均为聚集分布,但聚集度均有逐渐减弱的趋势。究其原因,可能是随着演替的进行,到生草沙丘阶段,由于前期钻蛀性害虫的危害取食,致使油蒿长势变弱,而野外调查发现,沙蒿大粒象和沙蒿木蠹蛾更易选择健康的油蒿进行危害; 同时还发现沙蒿木蠹蛾有转移危害的现象; 其次,由于油蒿属于半灌木,根从地表至地下15 cm处的平均粗度一般为1~3 cm,经过演替前2个阶段钻蛀性害虫的危害后,其干基部和根部大部分被蛀空,所剩的营养,很难满足钻蛀性害虫生长发育的需要,故油蒿钻蛀性害虫不得不选择周围长势较好的油蒿进行危害,使聚集度出现下降。
寄主的分布、寄主生境的空间结构以及天敌所感知的寄主丰富度的变化等均能影响到天敌的搜索行为(Roland et al., 1997)。研究蛀干害虫的分布特性及干扰因子对害虫生物学和生态学行为的影响机制,有利于制订合理的害虫防治措施。害虫的空间分布与种群的增殖密切相关,种群的空间格局反映种群的行为习性和栖境环境的影响,是种群有效利用资源的一个重要特征。研究种群空间格局可了解种群对环境变化表现出的适应性,并为种群抽样调查模型的建立提供基础(王小艺等,2005)。
对害虫具有良好控制作用的天敌生物必须对寄主害虫具有很强的搜索追踪和聚集攻击效应(邹运鼎,1997)。大量研究显示:很多害虫与其优势天敌种类在空间分布格局上密切相关,即优势天敌种类的分布型往往与其寄主害虫的分布规律一致(王小艺等,2005; 黄保宏等,2003; 陈向阳等,2006),除非受到外界因素(如喷洒杀虫剂)剧烈干扰而有所变化(郝树广等,2000),这是由优势天敌在时间和空间上对寄主害虫的强烈跟随效应造成的。本研究中寄生性天敌蒲螨和捕食性天敌斑翅山鹑啄痕的空间分布规律与其寄主或猎物的分布紧密相关,天敌的跟随作用明显。
本文研究了油蒿3个演替阶段中,4种油蒿钻蛀性害虫及寄生性天敌和捕食性天敌的空间分布情况。钻蛀性害虫可能多为聚集型分布,如郑元捷等(2006)研究表明杨树粒肩天牛(Apriona germari)幼虫在林间为聚集分布,陈向阳等(2006)发现松墨天牛(Monochamus alternatus)在纯林和混交林3种松树上的水平格局均为聚集分布,且其天敌花绒坚甲的水平格局亦为聚集分布,说明后者对前者在空间上有追随关系。
由于调查时主要以4种油蒿钻蛀性害虫的发生规律为主,因此,虽然4种油蒿钻蛀性害虫幼虫在各样地的抽样精度均符合要求,但因虫口密度不同,有些调查结果存在抽样精度偏低问题,这在对调查数据进行分析之前是难以察觉的。值得指出的是,评价昆虫种群空间格局的指标有很多,大致可以分为2类,一类指标明显与种群自身密度相关,如扩散系数C、平均拥挤度m*等,另一类则基本与种群密度无关,如k、m*/x等(刘树生等,1996)。有些指标的统计性质又是一致的,如C与I(丛生指标),k与Ca(聚集度指标),分析时不必一一罗列,如本研究中未选用I和Ca。此外,还应尽量选择与密度无关的指标,以区分生物因素和统计因素的作用。但每种指标经过计算后都将失去部分信息,不能完整反映真实的种群空间结构,因此在分析这些参数时应注意其局限性,一些密度相关指标尤其如此。
北京大学地理系, 中国科学院自然资源综合考察委员会, 中国科学院兰州沙漠研究所. 1983. 毛乌素沙区自然条件及其改良利用[M]. 北京: 科学出版社: 83-84.
|
陈向阳, 邹运鼎, 丁玉洲, 等. 2006. 松墨天牛及其天敌花绒坚甲种群的三维空间分布格局[J]. 应用生态学报, 17(8): 1547-1550. |
丁岩钦. 1994. 昆虫数学生态学[M]. 北京: 科学出版社.
|
郭柯. 2000. 毛乌素沙地油蒿群落的循环演替[J]. 植物生态学报, 24(2): 243-247. |
郝树广, 张孝羲, 程遐年. 2000. 稻田节肢动物群落优势功能集团的垂直分布、数量动态及天敌作用估计[J]. 应用生态学报, 14(3): 413-417. |
何志斌, 赵文智, 常学向, 等. 2004. 荒漠植被物种多样性对空间尺度的依赖[J]. 生态学报, 26(6): 1146-1149. |
黄保宏, 邹运鼎, 毕守东, 等. 2003. 朝鲜球坚蚧及黑缘红瓢虫空间格局的地统计学研究[J]. 应用生态学报, 14(3): 413-417. |
刘树生, 汪信庚, 吴晓晶, 等. 1996. 甘蓝和白菜上桃蚜种群的空间格局及其时序动态[J]. 昆虫学报, 39(2): 158-164. |
马立芹, 温俊宝, 许志春, 等. 2009. 寄生性蒲螨研究进展[J]. 昆虫知识, 46(3): 366-371. DOI:10.7679/j.issn.2095-1353.2009.068 |
王庆锁, 董学军, 陈旭东, 等. 1997. 油蒿群落不同演替阶段某些群落特征的研究[J]. 植物生态学报, 21(6): 531-538. |
王小艺, 杨忠岐. 2005. 白蜡窄吉丁幼虫及其天敌在空间格局上的关系[J]. 应用生态学报, 16(8): 1427-1431. |
王妍, 吴波, 卢琦, 等. 2006. 毛乌素沙地油蒿群落的物种组成与数量分类[J]. 北京林业大学学报, 28(S2): 52-56. |
邬祥光. 1985. 昆虫生态学的常用数学分析方法[M]. 北京: 农业出版社.
|
阎伟, 骆有庆, 宗世祥, 等. 2009b. 油蒿不同演替阶段钻蛀性害虫数量变动与环境的关系[J]. 林业科学, 45(6): 87-91. |
阎伟, 宗世祥, 骆有庆, 等. 2009a. 逐步回归模型在油蒿钻蛀性害虫预测中的应用[J]. 北京林业大学学报, 31(3): 140-144. |
杨洪晓, 张金屯, 吴波, 等. 2004. 油蒿(Artemisia ordosica)对半干旱区沙地生境的适应及其生态作用[M]. 北京师范大学学报: 自然科学版: 684-690.
|
余建英, 何旭宏. 2003. 数据统计分析与SPSS应用[M]. 北京: 人民邮电出版社.
|
张德魁, 王继和, 马全林, 等. 2007. 油蒿研究综述[J]. 草业科学, 24(8): 30-35. |
张军, 黄永梅, 焦会景, 等. 2007. 毛乌素沙地油蒿群落演替的生理生态学机制[J]. 中国沙漠, 27(6): 977-983. |
张新时. 1994. 毛乌素沙地的生态背景及其草地建设的原则与优化模式[J]. 植物生态学报, 18(1): 1-16. |
郑元捷, 陈顺立, 余培旺, 等. 2006. 杨树粒肩天牛幼虫的空间格局[J]. 华东昆虫学报, 15(3): 206-210. |
邹运鼎. 1997. 害虫管理中的天敌评价理论与应用[M]. 北京: 中国林业出版社: 50-64.
|
CCICCD(Chinese Committee for Implementing UN Convention to Combat Desertification). 1997. China Country Paper to Combat Desertification[M]. Beijing: China Forestry Publishing House: 40.
|
Roland J, Taylor P D. 1997. Insect parasitoid species respond to forest structure at different spatial scales[J]. Nature, 386: 710-713. DOI:10.1038/386710a0 |