 2011, Vol. 47
2011, Vol. 47文章信息
- 赵仲辉, 张利平, 康文星, 田大伦, 项文化, 闫文德, 彭长辉
- Zhao Zhonghui, Zhang Liping, Kang Wenxing, Tian Dalun, Xiang Wenhua, Yan Wende, Peng Changhui
- 湖南会同杉木人工林生态系统CO2通量特征
- Characteristics of CO2 flux in a Chinese Fir plantation ecosystem in Huitong County, Hunan Province
- 林业科学, 2011, 47(11): 6-12.
- Scientia Silvae Sinicae, 2011, 47(11): 6-12.
-
文章历史
- 收稿日期:2010-07-11
- 修回日期:2011-09-05
-
作者相关文章


2. 湖南会同杉木林生态系统国家野外科学观测研究站 会同 418307;
3. Institute of Environmental Science, University of Quebec at Montreal Montreal Quebec H3C3P8
2. Huitong National Research Station of Chinese Fir Plantation Ecosystem in Hunan Province Huitong 418307;
3. Institute of Environmental Science, University of Quebec at Montreal Montreal Quebec H3C3P8
“温室效应”是工业革命以来影响范围最广的生态环境问题之一(方精云,2000),CO2对全球变暖的贡献率达60%(Gifford,1994)。森林作为陆地生态系统的主体,它与大气之间的年碳交换量占陆地生态系统的90%(Winjum et al., 1993)。因此,量化森林生态系统的碳通量非常重要(Curtis et al., 2002)。
20世纪90年代以来,森林生态系统CO2通量研究取得了一些阶段性成果,如欧洲的森林生态系统大多表现为碳汇,碳吸收随纬度的增加而减少(Valentini et al., 2000); 光合有效辐射、空气温度、土壤水分含量是影响森林生态系统CO2通量日、季节变化的主要因素(Malhi et al., 1999; Lindroth et al., 1998; Dolman et al., 2002; Euskirchen et al., 2006); 干扰(Schulze et al., 1999)和林龄(Lloyd et al., 2001; Paw et al., 2004)也对森林生态系统CO2通量产生较大影响。ChinaFLUX的建立带动了中国区域碳通量观测的发展,促进了中国森林生态系统碳通量的研究。研究表明中国南方地区碳汇量占中国碳汇总量的65%以上,大面积的人工造林起了重要作用(Piao et al., 2009)。杉木(Cunninghamia lanceolata)是中国南方的主要造林树种,占中国人工林面积的30%,其总面积约921万hm2(雷加富,2005),可见杉木林在区域碳平衡中起着重要作用。但是,目前对杉木林生态系统碳循环过程的研究较少,主要采用生物量法或模型估算系统净生产力(谌小勇等,1996; 田大伦等,1998; Zhao et al., 2009; 侯振宏等,2009),其中生物量法只能计算出年碳平衡量,难以估算生态系统碳平衡的季节性变化及其与环境因子之间的关系,模型法需要确定大量的参数和模型验证,涡动相关法测定碳通量能弥补上述研究方法的不足,但目前还未见涡动相关法研究杉木林生态系统碳交换方面的报道。本研究利用2008年会同杉木人工林生态系统碳通量观测数据,研究杉木林生态系统碳吸存能力的变化规律及其影响因子,为进一步了解我国森林的碳汇功能提供科学依据。
1 研究区概况研究区位于湖南会同杉木林生态系统野外科学观测研究站(109°45′E,26°50′N),地貌为低山丘陵,海拔270~400 m。根据该站28年的气象观测资料统计,该区年平均气温17.1 ℃,极端最高气温39.1 ℃,极端最低气温-5.7 ℃,年均相对湿度87%,年降水量931.5~1 724.0 mm,年均降水量1 275.3 mm,属典型的中亚热带湿润季风气候; 地层属震旦纪板溪系灰绿色变质板页岩,土壤为山地黄壤,风化程度甚深,土层厚度80 cm以上。
碳通量观测塔安装在该站第2号集水区内东坡(坡度27°)的中上部,集水区面积2.0 hm2,以观测塔为中心,北边200 m以外依次为溪流、农田、村庄和公路,东边400 m以外为农田,南边300 m处有少量农田,西边数公里范围内均为杉木人工林。2008年全年及白天以西南风频率最高,分别为35.3%和22.2%,东北风频率最低,分别为3.1%和5.1%;夜间则以北风及西北风频率为高,2者共占47.9%,以南风频率最低,为3.2%。集水区内的杉木林是1996年营造的,原有的杉木林皆伐后经炼山、全垦整地,实生苗造林,2008年林分年龄为13年,平均胸径14.3 cm,平均树高12.6 m,平均密度1 920株·hm-2。林下植被较少,主要灌木为杜茎山(Maesa japonica)、柃木(Eurya japonica)、菝葜(Smilax china)、冬青(Ilex purpurea)和乌蕨(Stenoloma chusanum)等,主要草本植物有狗脊(Woodwardia japonica)、铁芒萁(Dicranopteris linearis)和华南毛蕨(Cyclosorus parasiticus)等(2008年11月调查数据)。
2 研究方法 2.1 测定方法涡动相关系统安装在高32.5 m的观测铁塔顶部的伸展臂(指向北偏东10°)上,由三维超声风速仪(CAST3,Campbell,USA)和开路CO2/H2O红外分析仪(LI—7500,Li—Cor,USA)组成,数据采样频率为10 Hz,数据传输给数据采集器(CR1000,Campbell,USA)进行计算处理,原始数据和30 min的通量计算结果贮存在PC卡上。
自动气象梯度观测系统包括5层(高分别为1.4,8.5,14.0,22.6和32.5 m)大气温湿度(HMP45C—L5932,Vaisala,Helsinki,Finland)、1层(16 m)净辐射(CNR1,Kipp & Zonen,Holland)和光合有效辐射(PAR—LIFE CAF1,Li—Cor,USA)、4层(土壤深度分别为5,15,30和60 cm)土壤温度(107—L,Campbell,USA)和土壤水分含量(CS616—L,Campbell,USA)及3层(土壤深度分别为10,20和45 cm)土壤热通量板(HFP01,Hukeflux,Netherlands)。各观测值通过数据采集器(CR1000,Campbell,USA)采集(采样频率为0.5 Hz),存储5 min平均值。
涡动相关法是通过测定垂直风速与大气中CO2浓度脉动量的协方差来确定植被与大气间CO2通量(于贵瑞等,2006),根据涡动相关理论,某物理量S的垂直湍流输送量FS可表示为FS=w′S′,w′为空气瞬时垂直风速脉动量,S′为物理量S的脉动量。
2.2 数据处理 2.2.1 通量的计算由于通量观测原始数据是基于理想条件的假设下所获得的,因此,只有对该原始数据经过一系列的校正和计算才可得到具有物理意义且能够用于表征生态过程的CO2通量值(于贵瑞等,2008)。一般来说,净生态系统CO2交换量(以后简记为NEE)NEE可用下式表示:NEE=Fc+Fc_WPL+Fa,Fc为经过坐标旋转后的观测值,Fc_WPL为WPL校正通量,Fa为冠层储存通量。NEE为正值表明生态系统向大气中释放CO2,即为碳源; NEE为负值表明生态系统从大气中吸收CO2,即为碳汇。
坐标轴旋转是为了消除因观测仪器安装存在倾斜或下垫面不平坦等因素对观测结果造成的影响,本研究采用3次坐标旋转; WPL校正是校正水热通量的传输对CO2通量所造成的影响; 在夜间,因大气层较稳定,土壤和植物呼吸所释放的CO2可能被储存在森林等高大植被冠层内,导致观测数据低于生态系统实际的碳交换量,所以必须进行冠层储存通量修正。在本研究中,探头高度以下的大气CO2储存项Fa利用CO2/H2O分析仪测定的CO2浓度进行计算(Greco et al., 1996; Pilegaard et al., 2001)。
2.2.2 数据的质量控制经过上述计算得到的CO2通量仍存在一些异常值,必须对其进行一系列的质量控制,主要包括降水同期数据剔除、阈值剔除、异常值剔除以及u*校正等。根据NEE数据范围,以0.2 mg CO2·m-2s-1为区间长度,分别统计白天和夜间的数据频率,确定白天NEE的阈值为[-1.2,0.4] mg CO2·m-2s-1(占白天样本数的93.3%),夜间NEE阈值为[-0.2,0.6] mg CO2 ·m-2s-1(占夜间样本数的85.5%)。考虑到测点地形的特殊性,异常值的剔除采用方差法和差分法(Papale et al., 2006)2种方法,以13天为时间步长,方差法中的阈值以3.6倍标准差为起点,以后每循环一次增加0.3,差分法的灵敏度取5.5。对于夜间弱湍流条件下造成的通量低估现象,通过剔除小于夜间摩擦速度的通量数据来解决,而临界摩擦速度是根据有效的夜间数据,用99%阈值法(Reichstein et al., 2005)确定其为0.1 m·s-1。考虑到植物的光合作用生理活动过程,当温度低于生物学零度时,光合作用过程停止,最后,将气温低于0 ℃及5 cm深处土壤温度低于5 ℃时对应的NEE<0的数据剔除。
2.2.3 数据的插补降雨雪期间探头探测路径受阻、停电、仪器维护及数据质量控制等会造成观测数据缺失(Pattey et al., 2002)。本研究的全部有效数据占所有观测数据的66.6%(其中白天80.3%,夜间54.1%),因此有必要对缺失数据进行插补。目前,常用的缺失数据插补方法有:平均日变化法、根据特定气象条件查表法和非线性回归法等。我们首先采用平均日变化法,对缺失数据用相邻几天同时刻数据的平均值进行替代,一般白天取14天的平均值、夜间取7天的平均值(Falge et al., 2001),插补过程中将白天数据和夜间数据分开处理。然后对未能插补的数据用非线性回归法插补,即对一定时段内有效的通量数据与气象数据建立回归统计模型,最后利用统计关系模型估算值来替换那些质量不可靠或缺测的通量数据。
2.2.4 通量的分离与统计数据处理的最终目的是获得不同时间尺度下完整的NEE、生态系统呼吸量(简记为RE)和总生态系统CO2交换量(简记为GEE)。本研究基于30 min的气象和通量数据,利用呼吸模型计算RE。由于夜间只有系统的呼吸作用,此时RE即为NEE,利用其与5 cm土壤温度的指数函数关系,得到呼吸模型中的参数,并将此参数推广到白天,计算出生态系统白天的RE,最后利用白天的NEE减去RE推算出白天的GEE。将获得的30 min通量数据累加求和,可以得到生态系统在日、月、年等不同时间尺度上的NEE,RE和GEE值(于贵瑞等,2006)。
3 结果与分析 3.1 主要气象因子变化2008年试验地主要气象因子日均值(日总量)的变化如图 1所示。32.5 m高处的气温和5 cm深处土壤温度月均值以7月份最高,分别为26.1和25.7 ℃,1月份最低,分别为1.9和4.1℃,1月中上旬和2月上旬正值2008年的冰灾时期,温度较低,4月则出现了2次高温天气,与7,8月日均气温接近; 光合有效辐射(简记为PAR)月总量最高值出现在7月,为38.1 mol·m-2,最小值出现在1月,为7.3 mol·m-2; 相对湿度月平均最高值出现在8月,为80.8%,最低值出现在2月,为70.8%;月降水总量8月份最多,为260.5 mm(多年平均最多在6月,为204.7 mm),12月份最少,仅11.8 mm,年降水总量为1 447.9 mm(多年平均为1 275.3 mm)。与多年平均情况相比,该年1,2月平均气温偏低2.5 ℃以上(2008年冰灾影响),11月降水是多年平均值的3.6倍,4月降水偏少,只有多年平均值的53.7%。

|
图 1 2008年主要气象因子年变化 Figure 1 Annual variation of main meteorological factors in 2008 |
全年白天的NEE与PAR关系见图 2,当PAR < 500 μmol·m-2s-1时,NEE随PAR的增大而明显增加,当PAR为500~1 500 μmol·m-2s-1时,NEE随PAR的增大而增加,但增幅减小,当PAR>1 500 μmol·m-2s-1后,NEE趋于稳定。

|
图 2 2008年白天净生态系统交换与光合有效辐射的关系 Figure 2 Relationship between net ecosystem exchange and photosynthetic active radiation during the day in 2008 |
用Michaelis-Menten模型(Michaelis et al., 1913)来拟合白天(PAR≥15 μmol·m-2s-1)NEE和PAR的关系:
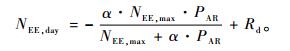
|
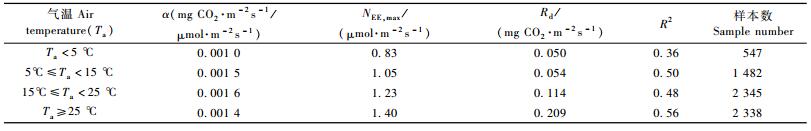
式中:NEE,day为白天NEE(mg CO2·m-2s-1); PAR为入射到冠层上的光合有效辐射(μmol·m-2s-1),α为表观初始光能利用率(mg CO2·m-2s-1/μmol·m-2s-1),NEE,max为光饱和时的NEE(mg CO2·m-2s-1) (即PAR→∞时NEE的渐进值),Rd(mg CO2·m-2s-1)为表观暗呼吸速率(即PAR→0时净生态系统交换值)。2008年不同气温(32.5 m高处)区间的拟合参数见表 1。由表 1可见,α随气温的增加而增大,在15~25℃时达到最大,之后,随气温上升而减小; NEE,max和Rd随气温的增加一直增大,这表明NEE对PAR的响应随气温的增加而增强。
|
|
土壤温度不仅影响根系呼吸,也会影响土壤微生物的活动。用每天夜间NEE求平均(由于所用数据为插补前的数据,故样本数不等,当样本数不够5个,则这天的数据不用),同时对每天夜间相应时次的不同深度的土壤温度求平均,发现夜间平均NEE与夜间5 cm深处土壤平均温度有显著的相关性(P < 0.05),即随着土壤温度升高,NEE呈指数增大(图 3),这与已有的一些研究结果基本一致(宋霞等,2004; 彭镇华等,2009)。

|
图 3 2008年夜间NEE与5 cm深处土壤温度的关系 Figure 3 Relationship between NEE and soil temperature at 5 cm depth during the night in 2008 |
2008年NEE4个季节代表月(1,4,7和10月)晴天平均日变化见图 4。1月(冬季)NEE在9:00—16:00为负(碳汇),其他时段为正(碳源),最小值出现在13:30(-0.37 mg CO2·m-2s-1),日较差为0.48 mg CO2·m-2s-1。4月(春季)NEE在7:30—17:00为负(碳汇),其他时段为正(碳源),最小值出现在11:30(-0.58mg CO2·m-2s-1),日较差为0.80 mg CO2·m-2s-1。7月(夏季)NEE在7:30—18:00为负(碳汇),其他时段为正(碳源),最小值出现在10:00(-0.78 mg CO2·m-2s-1),日较差为1.05 mg CO2·m-2s-1。10月(秋季)NEE在8:30—16:30为负(碳汇),其他时段为正(碳源),最小值出现在11:00(-0.62 mg CO2·m-2s-1),日较差为0.86 mg CO2·m-2s-1。

|
图 4 2008年会同杉木林NEE 4季代表月份晴天平均日变化 Figure 4 Mean diurnal variations of NEE in Chinese fir plantations ecosystem in four seasons, 2008 |
杉木林NEE的月变化见图 5。由图可见13年生杉木林NEE1—2月月总量为正值,表现为碳源,其余各月月总量都为负值,表现为碳汇。碳汇强度在生长旺季(5—8月)较大(-38.4~-53.0 g C·m-2),其他月份较小(-21.2~-12.4 g C·m-2),其中,最大碳汇出现在6月(-53.0 g C·m-2),最大碳源出现在1月(12.3 g C·m-2)。杉木林RE月变化为单峰型变化,7月最大(117.8 g C·m-2),2月最小(38.3 g C·m-2)。杉木林GEE的月变化特征与NEE的月变化基本一致,最小值出现在6月份(-157.7 g C·m-2),最大值出现在1月份(-26.5 g C·m-2)。2008年杉木林NEE,RE和GEE年总量分别为-255.3,958.3和-1 213.6 g C·m-2。

|
图 5 2008年RE,NEE和GEE的月总量变化 Figure 5 Annual variations of monthly accumulated RE, NEE and GEE in 2008 |
杉木林生态系统NEE与PAR、温度等气象因子存在明显的相关性。白天NEE随着PAR的不断增强而逐渐增大,但增大到一定程度则趋于稳定,用Michaelis-Menten模型模拟,效果较好,对不同温度水平下NEE与PAR的模拟结果表明,在同样的PAR条件下,随着温度升高,NEE增强。夜间平均NEE与5 cm深处土壤平均温度呈明显的指数关系,即随着土壤温度的升高而增强。
NEE具有明显的日变化和季节变化特征。在一天中,夜间NEE基本为正值且变化幅度小,白天NEE为负值且变化幅度大,NEE负值持续时间和NEE最小值出现的时间随季节的不同而不同,由冬季至春季至夏季,负值持续时间逐渐延长(分别为7.0,9.5和10.5 h),最小值出现的时间逐渐提前(分别为13:30,11:30和10:00),而由夏季至秋季至冬季,负值持续时间逐渐缩短(分别为10.5,8.0和7.0 h),最小值出现的时间逐渐推后(分别为10:00,11:00和13:30)。NEE是由光合作用和呼吸作用共同决定的,光合作用主要决定于PAR、空气温湿度和土壤水分含量等因素,而呼吸作用则主要决定于空气温度和土壤温度,这些因子综合作用决定了气孔导度的变化,从而影响NEE,例如在冬季的晴天,由于清晨气温和土壤温度比较低,日出以后一段时间,虽然PAR增强,植物也难以进行光合作用,从而使得NEE出现负值的时间推后,而在午后气温和土壤温度均达到最高值,这时PAR也较大,最有利于光合作用,从而使得NEE的最小值出现在午后; 而在夏季的晴天,日出时间提早,随着太阳高度的增大,PAR快速增长,很快就达到生态系统的光补偿点,从而使NEE出现负值的时间较早,随后,光合和呼吸作用均随辐射的增强和温度的升高而增强,如果前者快于后者,则使NEE降低,反之则使NEE升高,中午前后,PAR最强,气温又高,空气饱和差又大,植物呼吸加强,使叶片气孔关闭,反而使光合作用减弱,植物出现光合午休现象,从而使得NEE反而减小。在一年中,CO2通量月总量最小值出现在6月,为-53.0 g C·m-2,最大值出现在1月,为12.3 g C·m-2; 全年NEE为-255.3 g C·m-2,表现为碳汇,生态系统年呼吸量为958.3 g C·m-2,年总生态系统碳交换量为-1 213.6 g C·m-2。
中国通量网观测到的从北到南不同森林类型的NEE值中,除西双版纳总体表现为较弱碳汇外(张一平等,2006),其他森林生态系统NEE范围为-184.0~-645.0 g C·m-2 a-1(关德新等,2004; 刘允芬等,2004; 2006; 王春林等,2006),并随着纬度的增加而增大。本研究得到的湖南会同杉木人工林生态系统NEE(-255.3 g C·m-2a-1)远大于纬度相近的千烟洲(刘允芬等,2004; 2006)(-387.2~-645.0 g C·m-2 a-1),这种差异可能与林分、林龄、树高等有关,也可能与数据处理方法有关。在本研究中,夜间通量的u*剔除所找到临界摩擦速度小于0.1 m·s-1,故取0.1 m·s-1(Reichstein et al., 2005); 此外,考虑到当温度低于光合生理活动过程的生物学零度时,光合作用过程停止,但通量观测的结果却存在明显的碳汇,最后,将白天气温低于0 ℃及5 cm深处土壤温度低于5 ℃时对应的NEE<0的数据剔除并用夜间NEE与5 cm深处土壤温度的关系来插补,这2种考虑使得年NEE增大。
用生物量法和模型法得到会同杉木人工林的净初级生产力(简记为NPP)为344.8~690.5 g C·m-2a-1(谌小勇等,1996; 田大伦等,1998; Zhao et al., 2009),由于NPP并不等于NEE,如果要比较,则必须进行转换:根据NEE/NPP为-0.3~-0.4(Zhao et al., 2009),为简便起见,我们取NEE/NPP为-0.35,则用生物量和模型法测定的NPP换算得到的NEE为-120.7~-241.7 g C·m-2a-1; 根据NEE=0.90-0.56×NPP(Pregitzer et al., 2004),用生物量和模型法测定的NPP换算得到NEE为-192.2~-385.8 g C·m-2a-1。本研究用通量法估算出的13年生杉木人工林NEE(-255.3 g C·m-2a-1)与生物量法和模型法得到的结果基本一致。
谌小勇, 彭元英, 张昌建, 等. 1996. 亚热带两类森林群落产量结构及生产力的比较研究[J]. 中南林学院学报, 16(1): 1-7. |
方精云. 2000. 全球生态学:气候变化和生态响应[M]. 北京: 高等教育出版社.
|
关德新, 吴家兵, 于贵瑞, 等. 2004. 气象条件对长白山阔叶红松林CO2通量的影响[J]. 中国科学: D辑, 地球科学, 34(增刊Ⅱ): 103-108. |
侯振宏, 张小全, 徐德应, 等. 2009. 杉木人工林生物量和生产力研究[J]. 中国农学通报, 25(5): 97-103. |
雷加富. 2005. 中国森林资源[M]. 北京: 中国林业出版社.
|
刘允芬, 宋霞, 孙晓敏, 等. 2004. 千烟洲人工针叶林CO2通量季节变化及其环境因子的影响[J]. 中国科学: D辑, 地球科学, 34: 109-117. |
刘允芬, 于贵瑞, 温学发, 等. 2006. 千烟洲中亚热带人工林生态系统CO2通量的季节变异特征[J]. 中国科学: D辑, 地球科学, 36: 91-102. |
彭镇华, 王妍, 任海青, 等. 2009. 安庆杨树林生态系统碳通量及其影响因子研究[J]. 林业科学研究, 22(3): 237-242. |
宋霞, 刘允芬, 于贵瑞, 等. 2004. 红壤丘陵区人工林冬春时段碳、水、热通量的观测与分析[J]. 资源科学, 26(3): 97-104. |
田大伦, 盘宏华, 康文星, 等. 1998. 第二代杉木人工林生物量的研究[J]. 中南林学院学报, 18: 11-16. |
王春林, 于贵瑞, 周国逸, 等. 2006. 鼎湖山常绿针阔叶混交林CO2通量估算[J]. 中国科学: D辑, 地球科学, 36(增刊Ⅰ): 119-129. |
于贵瑞, 孙晓敏. 2006. 陆地生态系统通量观测的原理与方法[M]. 北京: 高等教育出社.
|
于贵瑞, 孙晓敏. 2008. 中国陆地生态系统碳通量观测技术及时空变化特征[M]. 北京: 科学出版.
|
张一平, 沙丽清, 于贵瑞, 等. 2006. 热带季节雨林碳通量年变化特征及影响因子初探[J]. 中国科学: D辑, 地球科学, 36(增刊Ⅰ): 139-152. |
Curtis P S, Hanson P J, Bolstad P, et al. 2002. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests[J]. Agricultural and Forest Meteorology, 113(1-4): 3-19. DOI:10.1016/S0168-1923(02)00099-0 |
Dolman A J, Moors E J, Elbers J A. 2002. The carbon uptake of a mid latitude pine forest growing on sandy soil[J]. Agricultural and Forest Meteorology, 111(3): 157-170. DOI:10.1016/S0168-1923(02)00024-2 |
Euskirchen E S, Pregitzer K S, Chen J Q, et al. 2006. Carbon fluxes in a young, naturally regenerating jack pine ecosystem[J]. Journal of geophysical research, 111: 1-13. |
Falge E, Baldocchi D, Olson R. 2001. Gap filling strategies for defensible annual sums of net ecosystem exchange[J]. Agricultural and Forest Meteorology, 107(1): 43-69. DOI:10.1016/S0168-1923(00)00225-2 |
Gifford D M. 1994. The global carbon cycle: a viewpoint on the missing sink[J]. Australian Journal of Plant Physiology, 21(1): 1-15. DOI:10.1071/PP9940001 |
Greco S, Baldocchi D D. 1996. Seasonal variations of CO2 and water vapor exchange rates over a temperate deciduous forest[J]. Global Change Biology, 2(3): 183-197. DOI:10.1111/gcb.1996.2.issue-3 |
Lindroth A, Grelle A, Moren A S. 1998. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity[J]. Global Change Biology, 4(4): 443-450. DOI:10.1046/j.1365-2486.1998.00165.x |
Lloyd J, Kolle O, Shibistova O, et al. 2001. The carbon balance of a central Siberian forest[J]. CGER. Proceedings of international workshop for advanced flux network and flux evaluation. Sapporo, Japan: Hokkaido University, 39: 39-45. |
Malhi Y, Baldocchi D D, Jarvis P G. 1999. The carbon balance of tropical, temperate and boreal forests[J]. Plant, Cell & Environment, 22(6): 715-740. |
Michaelis L, Menten M L. 1913. Die kinetik der invertinwirkung[J]. Biochemz, 49: 333. |
Papale D, Reichstein M, Aubinet M, et al. 2006. Towards a standardized processing of net ecosystem exchange measured with eddy covariance technique: algorithms and uncertainty estimation[J]. Biogeosciences, 3: 571-583. DOI:10.5194/bg-3-571-2006 |
Pattey E, Strachan I B, Desjardins R L, et al. 2002. Measuring nighttime CO2 flux over terrestrial ecosystems using eddy covariance and nocturnal boundary layer methods[J]. Agricultural and Forest Meteorology, 113(1-4): 145-158. DOI:10.1016/S0168-1923(02)00106-5 |
Paw U KT, Falk M, Suchanek T H, et al. 2004. Carbon dioxide exchange between an old-growth forest and the atmosphere[J]. Ecosystems, 7(5): 513-524. |
Piao S L, Fang J Y, Philippe C, et al. 2009. The carbon balance of terrestrial ecosystems in China[J]. Nature, 458: 1009-1013. DOI:10.1038/nature07944 |
Pilegaard K, Hummelshj P, Jensen N O, et al. 2001. Two years of continuous CO2 eddy-flux measurements over a Danish beech forest[J]. Agriculture Forestry Meteorology, 107(1): 29-41. DOI:10.1016/S0168-1923(00)00227-6 |
Pregitzer K S, Euskirchen E S. 2004. Carbon cycling and storage in world forests: biome patterns related to forest age[J]. Global Change Biology, 10(12): 2052-2077. DOI:10.1111/gcb.2004.10.issue-12 |
Reichstein M, Falge E, Baldocchi D, et al. 2005. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm[J]. Global Change Biology, 11(9): 1424-1439. DOI:10.1111/gcb.2005.11.issue-9 |
Schulze E D, Lloyd J, Kelliher J, et al. 1999. Productivity of forests in the Eurosiberian boreal region and their potential to act as a carbon sink asynthesis[J]. Global Change Biology, 5(6): 703-722. DOI:10.1046/j.1365-2486.1999.00266.x |
Valentini R, Matteucci G, Dolman A J, et al. 2000. Respiration as the main determinant of carbon balance in European forests[J]. Nature, 404: 861-865. DOI:10.1038/35009084 |
Winjum J K, Dixon R K, Schroeder P E. 1993. Forest management and carbon storage: an analysis of 12 key forest nations[J]. Water, Air & Soil Pollution, 70(1-4): 239-257. |
Zhao M F, Xiang W H, Peng C H, et al. 2009. Simulating age-related changes in carbon storage and allocation in a Chinese fir plantation growing in southern China using the 3-PG model[J]. Forest Ecology and Management, 257(6): 1520-1531. DOI:10.1016/j.foreco.2008.12.025 |