 2011, Vol. 47
2011, Vol. 47文章信息
- 卜庆梅, 柏新富, 朱建军
- Bu Qingmei, Bai Xinfu, Zhu Jianjun
- 4种园林树木叶绿素荧光参数对脱水的响应及耐旱特性分析
- Responses of Chlorophyll Fluorescence Parameters in Four Garden Tree Species to Desiccation Stress and Analysis of Their Drought Tolerance
- 林业科学, 2011, 47(10): 37-43.
- Scientia Silvae Sinicae, 2011, 47(10): 37-43.
-
文章历史
- 收稿日期:2010-09-30
- 修回日期:2011-03-30
-
作者相关文章



水分是植物生长和生存繁殖最为关键的生态因子,干旱则是影响植物生长最大的环境因素之一。干旱胁迫常影响植物的生长发育、造成光合产物积累减少(Li et al., 2006;邱念伟等,2008)。有关植物耐旱性能的分析研究已有很多报道,但相关研究多以盆栽试验为依据,或是对环境水分状况变化对植物生长、形态及生理生化变化状况进行观察分析。盆栽试验难以反映多年生树木实际生长状况,以环境水分变化为依据也不能真实显示植物体内的实际水分状态。而关于植物在不同体内水分状况条件下生理生态特征变化的研究则较少,且均限于荒漠植物(施征等,2008;柏新富等,2008)。
在干旱胁迫下,植物叶片脱水导致气孔关闭,从而限制光合作用;在叶片严重脱水时就会破坏光合机构(Georgieva et al., 2005)。叶绿素荧光测定和分析技术因其方便快速、灵敏和无损伤地反映逆境胁迫下光合机构功能的变化而在植物环境生理研究中得到广泛应用(李鹏民等,2005;Hsu,2007)。园林植物在美化和改善城镇及道路环境中具有非常重要的作用,研究园林植物光合性能对体内水分状况的响应, 不仅能够分析这些植物耐旱能力的强弱,为绿化环境适地适树提供可靠的依据,而且有助于了解植物抵御干旱和水分胁迫的内在机理。本试验以我国北方常见的园林树木:国槐(Sophora japonica)、加杨(Populus ×canadensis)、银杏(Ginkgo biloba)和黑松(Pinus thunbergii)为材料,通过实地采样,对其叶片叶绿素荧光随体内水分变化的指标进行测定,以分析、比较这4种植物的耐旱特点,为造林绿化树种的选择、苗木的选育提供参考。
1 材料与方法 1.1 试验材料国槐、加杨、银杏和黑松均采自鲁东大学校园内。选立地条件基本一致,长势旺盛的向阳面枝条为材料。剪取枝条顶端约20 cm,称鲜质量后浸入水中3.5 h,使其水分饱和,取出吸干外表水分,称量饱和鲜质量。随后平铺于室内纸板上,在光照强度200 μmol·m-2s-1、温度25~27 ℃、相对湿度59%~60%条件下脱水,每隔30 min(国槐为15 min)称质量1次并测定记录叶片的叶绿素荧光参数。
1.2 试验方法 1.2.1 叶绿素荧光参数的测定叶绿素荧光参数用便携式植物效率分析仪(Handy-PEA,Hansatech,英国)测定。激发光强为3 000 μmol·m-2s-1,暗适应时间为15 min,记录时间1 s。荧光参数Fo, Fm, Fv, Fv/Fm, Fv/Fo, PI等用Handy PEA软件直接从测定结果中导出,RC/CSo, ABS/RC, Wk, ETo /ABS和Sm等参照李鹏民等(2005)的方法计算。所有测定均重复测定10个叶片(取自5个植株的5个枝条,每个枝条以从顶端数第1, 2个成熟叶片为测定对象),结果以平均数记。为便于不同植物之间的比较,各参数标准化后再作图、分析。
1.2.2 植物体相对含水量的测定相对含水量参照孙山等(2008)的方法:相对含水量RWC =[(鲜质量-干质量)/(饱和鲜质量-干质量)]×100%
2 结果与分析 2.1 植株相对含水量随脱水时间的变化通过对国槐、加杨、银杏和黑松4种园林植物叶片相对含水量(RWC)随脱水时间变化的测定发现:虽然4种植物的RWC均随脱水时间延长而降低,但其叶片保水能力却存在明显差异(图 1)。国槐在离体2.5 h后,叶片的相对含水量(RWC)已低于10%;加杨则在离体7.5 h后,RWC才开始低于10%;而银杏在离体11.5 h后,RWC为17.9%;黑松则在离体36 h后仍维持高达49.2%的RWC。说明黑松和银杏能更好地保持体内水分以适应干旱和空气干燥环境。

|
图 1 4种树木相对含水量随脱水时间的变化 Figure 1 Changes of RWC with desiccation time in four tree species |
国槐、加杨、银杏和黑松脱水过程中快速叶绿素荧光动力学曲线变化如图 2。可以看出:所测定的4种树木在脱水情况下,叶绿素荧光的强度和动力学曲线的形状都发生大幅度的变化;同时,不同植物在相同脱水时间的叶绿素荧光的强度和曲线变化幅度又存在较大差异,比如,在脱水4 h的叶绿素荧光动力学曲线,国槐已趋于水平,加杨和银杏虽有较大变化但曲线的O, J, I, P点仍然非常明显,而黑松则变化较小;加杨和银杏在脱水7 h时,叶绿素荧光动力学曲线趋于水平,黑松在脱水36 h后才趋于水平。表明不同植物在脱水过程中其光合系统性能的变化复杂且各异。

|
图 2 4种树木不同脱水时间的快速叶绿素荧光动力学曲线 Figure 2 The rapid fluorescence induction kinetics of four tree species during desiccation |
Fv/Fm表示PSⅡ最大光化学效率,是反映光合机构生理状态和光合作用受影响程度的重要指标之一(胡学华等,2007;李志军等,2009)。在正常条件下,黑松、银杏、加杨和国槐叶片的Fv/Fm分别为0.858, 0.814, 0.856和0.855,除银杏稍低外,其他三者基本相同(图 3)。随着脱水时间的延长,国槐叶片的Fv/Fm快速降低,加杨和银杏也先后大幅下降,三者分别在3.5, 7和10 h以内降至0.1左右;而黑松则下降缓慢,在35~36 h才降至0.1左右。然而以相对含水量为标准进行比较,则黑松、银杏、加杨和国槐叶片的Fv/Fm分别在RWC约为70%, 50%, 40%和20%时出现快速下降,在RWC约为50%, 30%, 10%和7%时降至0.1左右。由此可见:黑松虽然抗脱水能力很强,但其光合系统对脱水却非常敏感;而国槐则正好相反。

|
图 3 4种树木Fv/Fm随脱水时间和相对含水量的变化 Figure 3 The changes of Fv/Fm with desiccation time and RWC in four tree species during dehydration |
在荧光诱导动力学参数的测定中,经暗适应的叶片可变荧光(Fv)与初始荧光(Fo)的比值可代表PSⅡ的潜在活性(张守仁,1999;赵丽英等,2005)。对4种树木的测定计算发现,随着脱水胁迫程度的增加,Fv/Fo呈逐渐降低趋势(图 4),其中黑松下降速度最快,银杏次之,加杨第三,国槐最慢,表明国槐光合系统对脱水有更高的稳定性。

|
图 4 4种树木Fv/Fo随相对含水量的变化 Figure 4 Changes of Fv/Fo with RWC in fore tree species |
RC/CSo表示单位面积有活性反应中心的数量、ABS/RC反映单位反应中心吸收的光能(李鹏民等,2005)。对4种园林树木的RC/CSo和ABS/RC随叶片相对含水量(RWC)的降低而变化的分析显示:脱水初期RC/CSo和ABS/RC均无明显变化,黑松在脱水至RWC为60%时出现RC/CSo的迅速下降和ABS/RC快速上升;银杏、加杨和国槐分别在脱水至RWC为50%,30%和10%时才出现RC/CSo的迅速下降和ABS/RC的快速上升(图 4,5)。RC/CSo的下降表明单位面积有活性反应中心数量的减少,ABS/RC上升则说明每个有活性反应中心吸收的光能增加(李鹏民等,2005)。两者的变化规律显示:在脱水过程中光合反应中心部分受到损伤、有活性的反应中心数目下降,使剩余的有活性的反应中心的负担加重,迫使剩余的有活性的反应中心效率提高。4种树木RC/CSo和ABS/RC的不同变化表明国槐、加杨、银杏和黑松光合反应中心对脱水胁迫的稳定性依次降低。

|
图 5 4种树木ABS/RC和RC/CSo随相对含水量的变化 Figure 5 Changes of ABS/RC and RC/CSo with RWC in four tree species |
在叶绿素荧光诱导曲线中K点的出现可以作为放氧复合体受伤害的一个特殊标记(李鹏民等,2005)。Wk为K点相对可变荧光的标准化,表示K点相对值的大小,Wk越大,放氧复合体受到的伤害越严重(孙山等,2008)。对黑松、银杏、加杨和国槐叶片Wk的计算发现,在RWC从100%降至40%时,Wk虽略有升高的趋向,但变化并不明显(图 6)。这说明脱水对黑松、银杏、加杨和国槐叶片放氧复合体的影响不大。

|
图 6 4种树木Wk随相对含水量的变化 Figure 6 Changes of Wk with RWC in four tree species |
ETo/ABS反映了反应中心吸收的光能用于电子传递的量子产额,其值越大说明PSⅡ具有高的电子传递活性(衣艳君等,2007)。Sm为标准化后的O-J-I-P荧光诱导曲线、荧光强度F=Fm及y轴之间的面积,Sm反映PSⅡ反应中心受体侧PQ库的大小,当电子从QA-进入电子传递链越多,Sm的值也越大(李鹏民等,2005)。脱水胁迫引起4种绿化树木叶片ETo /ABS和Sm的降低(图 7),说明脱水导致PSⅡ电子传递活性降低和受体侧PQ库的减小。但不同植物的表现存在明显差别,随着相对含水量的减少,黑松和银杏的ETo /ABS和Sm快速降低,而加杨和国槐的下降速度相对较慢;黑松和银杏在RWC降至50%~60%时,光合电子传递效率已显著降低,而加杨和国槐在RWC20%~30%时仍能维持一定的光合电子传递潜力。

|
图 7 4种树木ETo /ABS和Sm随相对含水量的变化 Figure 7 Changes of ETo /ABS and Sm with RWC in four tree species |
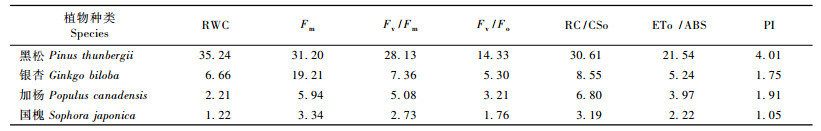
为了探明4种树木荧光参数变化与脱水时间的关系,对RWC降至初始值的50%和Fo, Fm, Fv/Fm, Fv/Fo, RC/CSo, ETo /ABS, PI等主要荧光参数变化至初始值的50%(其中Fo为上升,其他为下降)时所经历的脱水时间进行计算,结果见表 1。各参数变化至初始值的50%所经历的脱水时间以黑松最长、银杏较长、杨树较短、国槐最短,表明4种植物主要荧光参数的稳定性相对于脱水时间存在明显不同,即黑松最为稳定、银杏次之、加杨再次、国槐最不稳定。在各荧光参数中,PI变化最快,在较短的时间内就下降至初始值的50%,表现出对光合机构变化的高度灵敏性,这也与已有的研究结果相一致(Van Heerden et al., 2004; 李鹏民等,2005)。
|
|
光合作用是构成植物生物量的基础,也是植物对干旱最为敏感的生理过程之一(刘慧民等,2010)。有研究证明:轻度干旱通过气孔限制(减少CO2的进入)而使光合速率下降(王玉珏等,2010);持续的干旱则导致光系统Ⅱ(PSⅡ)活性降低和激发能的上升,活性氧浓度升高,伤害放氧复合体(OEC)、PSⅡ反应中心和电子传递链,引起PSⅠ/PSⅡ失衡、电子传递功能下降,并最终导致光合系统的损伤(Müller et al., 2001;李耕等,2009)。作为一种研究植物光合功能的快速、无损伤探针,叶绿素荧光动力学技术在测定叶片光合作用过程中光系统对光能的吸收、传递、耗散和分配等方面具有独特的优势,与表观性的气体交换指标相比,叶绿素荧光参数更具有反映内在性的特点(Genty et al., 1989; Scheriber et al., 1994)。任何影响光合作用的环境因素均会导致叶绿素荧光特性的改变(Wright et al., 2009)。在本试验中,随着脱水时间的延长,水分胁迫程度加剧,Fv/Fm呈明显的下降趋势;同时,反映PSⅡ潜在活性的Fv/Fo、反映PSⅡ反映中心状态的RC/CSo和ABS/RC以及反映PSⅡ受体侧电子传递情况的ETo /ABS和Sm等均随叶片相对含水量(RWC)的减少(水分胁迫程度加剧)而下降。但4种植物间也存在显著的不同,以RWC为基础,随RWC的降低,黑松的各种荧光参数下降最快,表现出对体内水分匮缺非常敏感;银杏、加杨和国槐的敏感性则依次降低。然而,如果以脱水时间为基础,随脱水时间的延长,国槐的各种荧光参数下降最快,表现出极弱的抗脱水能力;加杨和银杏比国槐稍慢,而黑松的各种荧光参数随脱水时间的延长变化非常缓慢,其Fv/Fm在脱水28.13 h的下降比例只相当于国槐脱水2.73 h的大小,显示出极强的抗脱水能力。
植物的形态结构与自然生境有关,其结构变化与机能的统一是植物适应生长环境的生物学基础(汪贵斌等,2008)。不同种类的植物在长期进化中形成了形态结构和生理生化方面不同的耐环境胁迫途径,叶片角质层厚、气孔下陷、栅栏组织/海绵组织比值高、表皮毛发达等都是植物耐水分亏缺的重要指标(李正理,1981)。黑松具有针状叶,针叶表面具有较厚的角质层和蜡质层,气孔下陷、气孔的前腔壁覆盖着大量的蜡质(邵邻相等,2005);银杏叶表面具明显的角质层和蜡质层,气孔保卫细胞下陷且围绕的4~7个副卫细胞顶上突起覆盖了气孔(李正理等,1989;陈立群等,2004);加杨叶片也具有蜡质层和中等或较厚的角质层(樊汝汶,1985);国槐虽然也具有一定的耐旱能力(柴胜丰等,2008),但其叶片防止水分散失的结构特征并无特别之处。从叶片形态特征分析,在本试验所研究的4种树木中,抗脱水能力以黑松最强、银杏较强、加杨相对较弱、国槐最弱,与相对含水量以及叶绿素荧光特性随脱水时间变化的结果完全一致。但是,如果以相对含水量(RWC)变化为横坐标,把荧光参数突然大幅降低的相对含水量称为相对含水量变化的阈值(衣艳君等,2007),则发现黑松在RWC降至60%~70%时,叶片的最大光化学效率(Fv/Fm)、反应中心电子传递效率(ETo /ABS)和有活性反应中心数量(RC/CSo)等反映其叶片光合性能的重要参数均出现快速大幅下降,即相对含水量的阈值为60%~70%;而银杏、加杨和国槐这些参数快速降低的RWC阈值分别为40%~50%,30%~40%和10%~20%,即黑松对失水最为敏感,银杏次之,加杨再次,国槐最不敏感。由此可见:黑松抗脱水能力很强但其光合系统对失水非常敏感,主要是通过限制水分丧失来抵抗干旱;国槐抗脱水能力很弱但其光合系统却能在失水较多的情况下保持较高的性能,忍耐脱水的能力较强;银杏和加杨则居两者之间。这一结果与李吉跃等(1993)对松类耐旱机理的研究以及何茜等(2007)对国槐蒸腾耗水的研究结果一致。
综合4种植物叶片形态结构特点和本试验结果可知:国槐和加杨虽然具有一定的耐脱水能力,但由于失水较快,耗水量大,在缺水地区不适宜大面积栽植,特别是国槐只能用于水分条件较好的立地条件下造林;黑松则具有抗脱水能力强、失水慢、耗水少的特点,但对脱水的耐力不强,可在半干旱区和干燥的半湿润区大面积造林种植;银杏耐旱能力强于杨树但远不及黑松,可以在干燥的半湿润区和湿润区造林种植。
柏新富, 朱建军, 赵爱芬, 等. 2008. 几种荒漠植物对干旱过程的生理适应性比较[J]. 应用与环境生物学报, 14(6): 763-768. |
柴胜丰, 蒋运生, 韦霄, 等. 2008. 干旱胁迫对槐树幼苗生长和生理生态特征的影响[J]. 浙江林业科技, 28(2): 43-47. |
陈立群, 李承森. 2004. 银杏(Ginkgo biloba)叶表皮特征及其气孔的发育[J]. 植物研究, 24(4): 417-422. |
樊汝汶. 1985. 黑杨派和青杨派叶片的比较解剖[J]. 南京林学院学报, 9(2): 40-46. |
何茜, 李吉跃, 齐涛. 2007. 植物生长调节剂对国槐蒸腾耗水的影响[J]. 北京林业大学学报, 29(1): 74-78. |
胡学华, 蒲光兰, 肖千文, 等. 2007. 水分胁迫下李树叶绿素荧光动力学特性研究[J]. 中国生态农业学报, 15(1): 75-77. |
李耕, 高辉远, 赵斌, 等. 2009. 灌浆期干旱胁迫对玉米叶片光系统活性的影响[J]. 作物学报, 35(10): 1916-1922. |
李吉跃, 张建国. 1993. 北方主要造林树种耐旱机理及其分类模型的研究[J]. 北京林业大学学报, 15(3): 1-11. |
李鹏民, 高辉远, StrasserR J. 2005. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用[J]. 植物生理学与分子生物学学报, 31(6): 559-566. |
李正理. 1981. 旱生植物的形态和结构[J]. 生物学通报, (4): 9-12. |
李正理, 贺晓, 徐炳文. 1989. 银杏叶表皮结构[J]. 植物学报, 31(6): 427-431. |
李志军, 罗青红, 伍维模, 等. 2009. 干旱胁迫对胡杨和灰叶胡杨光合作用及叶绿素荧光特性的影响[J]. 干旱区研究, 26(1): 45-52. |
刘慧民, 车艳双, 车代弟. 2010. 干旱胁迫对华北绣线菊和金山绣线菊光合能力的影响[J]. 应用生态学报, 21(8): 2004-2009. |
邱念伟, 杜斐, 郝爽, 等. 2008. 叶片喷施甜菜碱在烟草叶片保水和耐脱水中的作用[J]. 中国农业科学, 41(10): 3363-3370. DOI:10.3864/j.issn.0578-1752.2008.10.059 |
邵邻相, 张凤娟. 2005. 6种松科植物叶表皮的扫描电镜观察[J]. 植物研究, 25(3): 281-285. |
施征, 史胜青, 肖文发, 等. 2008. 脱水胁迫对梭梭和胡杨苗叶绿素荧光特性的影响[J]. 林业科学研究, 21(4): 566-570. |
孙山, 王少敏, 王家喜, 等. 2008. 黑暗中脱水对'金太阳'杏离体叶片PSⅠ和PSⅡ功能的影响[J]. 园艺学报, 35(1): 1-6. |
汪贵斌, 曹福亮, 景茂, 等. 2008. 水分胁迫对银杏叶片叶肉细胞超微结构的影响[J]. 南京林业大学学报:自然科学版, 32(5): 65-70. |
王玉珏, 付秋实, 郑禾, 等. 2010. 干旱胁迫对黄瓜幼苗生长、光合生理及气孔特征的影响[J]. 中国农业大学学报, 15(5): 12-18. |
衣艳君, 刘家尧. 2007. 毛尖紫萼藓PS Ⅱ光化学效率对脱水和复水的响应[J]. 生态学报, 27(12): 5238-5244. DOI:10.3321/j.issn:1000-0933.2007.12.034 |
张守仁. 1999. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 16(4): 444-448. |
赵丽英, 邓西平, 山仑. 2005. 渗透胁迫对小麦幼苗叶绿素荧光参数的影响[J]. 应用生态学报, 16(7): 1261-1264. |
Genty B, Brlantals J M, Baker N R. 1989. The relationship between quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J]. Biochimica Biophysica Acta, 990(1): 87-92. DOI:10.1016/S0304-4165(89)80016-9 |
Georgieva K, Maslenkova L, Peeva V, et al. 2005. Comparative study on the changes in photosynthetic activity of the homoiochlorophyllous desiccation-tolerant Haberlen rhodopensis and desiccation-sensitive spinach leaves during desiccation and rehydration[J]. Photosynthesis Research, 85(2): 191-203. DOI:10.1007/s11120-005-2440-0 |
Hsu B D. 2007. On the possibility of using a chlorophyll fluorescence parameter as an indirect indicator for the growth of Phalaenopsis seedlings[J]. Plant Science, 172(3): 604-608. DOI:10.1016/j.plantsci.2006.11.006 |
Li R H, Guo P P, Baumz M, et al. 2006. Evaluation of chlorophyll content and fluorescence parameters as indicators of drought tolerance in barley[J]. Agricultural Sciences in China, 5(10): 751-757. DOI:10.1016/S1671-2927(06)60120-X |
Müller P, Li X P, Niyogi K K. 2001. Non-photochemical quenching: A response to excess light energy[J]. Plant Physiology, 125(4): 1558-1566. DOI:10.1104/pp.125.4.1558 |
Scheriber U, Bilger W, Neubauer C. 1994. Chlorophyll fluorescence as a non-destructive indicator for rapid assessment of in vivo photosynthesis[J]. Ecological Studies, 100(1): 49-70. |
Van Heerden P D R, Strasser R J, Krüger G H J. 2004. Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics[J]. Physiologia Plantarum, 121(2): 239-249. DOI:10.1111/ppl.2004.121.issue-2 |
Wright H, DeLong J, Lada R, et al. 2009. The relationship between water status and chlorophyll a fluorescence in grapes (Vitis spp)[J]. Postharvest Biology and Technology, 51(2): 193-199. DOI:10.1016/j.postharvbio.2008.07.004 |