 2011, Vol. 47
2011, Vol. 47文章信息
- 时忠杰, 徐大平, 高吉喜, 宋爱云, 于春堂, 张宁南, 胡哲森
- Shi Zhongjie, Xu Daping, Gao Jixic, Song Aiyun, Yu Chuntang, Zhang Ningnan, Hu Zhesen
- 海南岛尾细桉人工林碳贮量及其分布
- Carbon Storage and Its Distribution of Eucalyptus urophylla×E.tereticornis Plantations in Hainan Island, Southern China
- 林业科学, 2011, 47(10): 21-28.
- Scientia Silvae Sinicae, 2011, 47(10): 21-28.
-
文章历史
- 收稿日期:2010-06-08
- 修回日期:2011-09-01
-
作者相关文章



2. 中国林业科学研究院热带林业研究所 广州 510520;
3. 环境保护部南京环境科学研究所 南京 210042;
4. 中国生态学会 北京 100085;
5. 北京能环科技发展中心 北京 100025;
6. 福建农林大学林学院 福州 350002
2. Research Institute of Tropical Forestry, CAF Guangzhou 510520;
3. Nanjing Institute of Environmental Sciences, Ministry of Environmental Protection of the People's Republic of China Nanjing 210042;
4. Ecological Society of China Beijing 100085;
5. Energy and Environmental Development Research Center Beijing 100025;
6. College of Forestry, Fujian University of Agriculture and Forestry Fuzhou 350002
随着工业化进程加快,大量化石燃料燃烧和土地利用方式改变等人为因素导致大气CO2浓度不断升高,CO2浓度升高是全球气候变化的最主要驱动力之一,气候变暖成为全球关注的重大科学问题(Marland et al., 2003;West et al., 2002;唐红侠等,2009)。人工林的固碳功能被认为是减缓全球气候变化的一种最有希望的选择,为减缓全球气候变化,1997年的《联合国气候变化框架公约》提出可以通过陆地生态系统有效管理提高固碳潜力,以达到减排的目的。研究人工林碳贮量及其固碳功能,对于评价森林生态系统碳汇潜力,提供碳汇林业对策具有重要意义。
桉树(Eucalyptus)原产于澳大利亚、印度尼西亚等热带地区,是十分优良的短轮伐期纸浆和用材树种,目前在我国华南地区发展十分迅速,面积已经超过300万hm2(中国工程院桉树项目组,2009)。桉树人工林对生态环境的影响特别引人关注,有关桉树人工林生物多样性、水分消耗及土壤肥力等的研究报道较多(茶正早等,1999;陈秋波,2001),但对其固碳功能的研究还不多。桉树人工林具有速生性和高生产力,在固碳潜力方面具有很大优势,因此对桉树人工林合理发展和有效管理,发挥其固碳效益具有重要意义。本研究基于海南西部沿海台地区、北部平原区、东部沿海台地区和中部山地区共18个调查区54个尾细桉(Eucalyptus urophylla×E. tereticornis)人工林样地调查数据,分析海南尾细桉人工林的生物量、碳贮量、固碳能力及其区域空间分布特征,为正确认识尾细桉人工林的碳汇功能及其对减缓气候变化的贡献提供依据。
1 研究区概况海南岛地处热带北缘(108°37′—111°03′E,18°10′—20°10′N),地形中部高,四周低,属热带季风气候,年平均气温22~26 ℃,≥10 ℃年积温8 200 ℃,最冷月1和2月温度仍达16~21℃,年光照1 750~2 650 h,年均降雨量1 639 mm,降水分布不均,东湿西干明显,中部的琼中山区雨量最大,以西部的东方市雨量最小,降水季节差异明显,5—10月为多雨季,降雨量达1 500 mm左右,占全年总降雨量的70%~90%,降水以台风雨和锋面雨为主,11月至翌年4月为旱季,降雨量仅占全年的10%~30%。
海南植被类型多样,自然植被类型主要有热带雨林、热带季雨林、山地雨林、针叶林和红树林等,占全岛森林面积的37.1%,人工林主要有橡胶(Hevea brasiliensis)林、桉树(Eucalyptus spp.)林、马占相思(Acacia mangium)林和芒果(Mangifera indica)林。土壤以砖红壤和山地黄壤为主,以砖红壤面积最大。
桉树人工林是20世纪80年代以后开始迅速发展起来的。目前全岛种植的桉树人工林约33.33万hm2(中国工程院项目组,2009),占全岛森林面积的10.1%,主要分布在海口、定安、澄迈、临高、儋州、白沙和东方等县市的沿海台地,东部的琼海、万宁也有一定的种植面积。
尾细桉人工林林下灌木层不发达,以草本植物为主,林下植物主要有山黄麻(Trema tomentosa)、桃金娘(Rhodomyrtus tomentosa)、白楸(Mallotus paniculatus)、浓子茉莉(Fagerlindia scandens)、毛蔓豆(Calopogonium mucunoides)、凤头黍(Acroceras munroanum)和假臭草(Eupatorium catarium)等。
2 研究方法2008年4—6月,根据土壤、气候等特征,在海南东部、西部、北部和中部选择18个调查区,调查区概况见表 1,每个调查区设置3块30 m×30 m样地,共计54块样地,样地多位于平坦地形,林分郁闭度0.3~0.7,林龄0.1~7.5年,桉树品种为尾细桉。
|
|
在样地内进行每木调查,测定并记录所有林木的胸径和树高,每隔2 cm为一个径级,共8个径级(0~2, 2~4, 4~6, 6~8, 8~10, 10~12, 12~14和14~16 cm),选取各径级标准木2~3株伐倒,共伐21株。对树干采用“分层切割法”(马明东等,2007;张治军等,2009),每1 m为一区分段,分树干、树皮、树枝及树叶;地下部分采用全根挖掘法,区分根桩、粗根(根系直径≥2 cm)、中根(1~2 cm)和细根( < 1 cm)。分别称各组分的鲜质量,对各组分取样,重复3次,并在80 ℃下烘干至恒质量,计算各部分的含水率和干质量,然后利用相对生长法估算全林分的生物量。利用树木胸径与各器官生物量数据,按W=a(Db)拟合尾细桉单株各器官的生物量与胸径的关系,树干生物量Wtrunk、树皮生物量Wbark、树叶生物量Wleaf、树枝生物量Wbranch和树根生物量Wroot拟合方程分别为:Wtrunk=0.080 9D2 317; Wbark=0.014 4D2.379 6; Wleaf=0.000 7D2.880 9; Wbranch=0.009 7D2.785 8和Wroot=0.014 5D2.746 4。
在每个样地内随机设置4块2 m×2 m小样方,采用样方收获法测定灌草层地上部分和地下部分的鲜质量,并在每块样方内分别选取3个重复的地上和地下部分样品,带回室内80 ℃烘箱内烘干至恒质量,测定含水率计算灌草层地上部分和地下部分的生物量;在每个样地内随机设置5块1 m×1 m小样方,收集全部凋落物,带回室内直接80 ℃烘干并测定其干质量,计算枯落物现存量。
由于尾细桉人工林树叶更新较快,直接将尾细桉人工林的叶现存量作为尾细桉人工林的年净生产量:
式中:ΔW为年净生产量(t·hm-2 a-1);Wm为m年时单位面积现存生物量(t·hm-2),Wm-n为m-n年时单位面积现存生物量(t·hm-2)。
由于尾细桉林下植被以草本植物为主,灌木相对较少,因此将林下植被生物量作为林下植被的年生产量。
在测定生物量的同时,采集乔木层各器官(叶、侧枝、主枝、树干、树皮、根桩、粗根、中根、细根)、灌草层、凋落物层样品,经烘干、粉碎、过筛后待测;在每个样地内选择2~3个代表性采样点挖土壤剖面,分层(0~20,20~40,40~60,60~80和80~100 cm)采集土壤样品,并将每块样地按土层分别混合后取部分土样,带回室内自然风干、磨碎待测;土壤密度通过环刀法测定,每个土壤剖面每层3个重复。植物和土壤样品碳元素含量均采用重铬酸钾氧化-外加热法测定(中国农业化学专业委员会,1983)。
尾细桉不同器官(叶、枝、干、皮、根)生物量、林下植被层生物量、凋落物现存量与各自部分碳含量的乘积之和为碳贮量,土壤各层的密度、体积与有机碳含量的乘积之和为地下土壤碳贮量,生物质部分和土壤部分之和为尾细桉人工林生态系统的碳贮量。
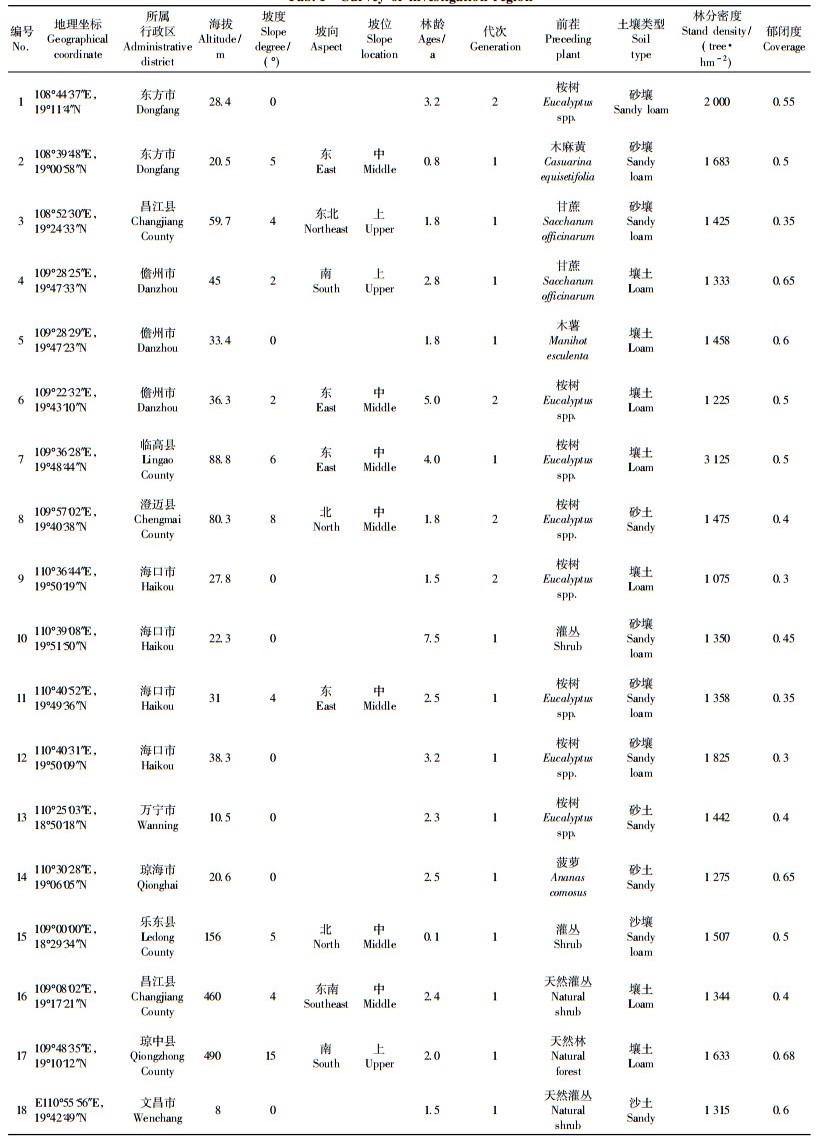
3 结果与分析 3.1 尾细桉人工林各组分碳含量由表 2可见,尾细桉林分乔木层各器官碳含量为438.12~501.34 g·kg-1,表现为树叶>树干>树根>树枝>树皮,不同器官碳含量变异系数为1.57%~5.65%。林下植被主要为矮灌木和草本植物,其碳含量平均为475.32 g·kg-1,凋落物层碳含量平均为479.61 g·kg-1。
|
|
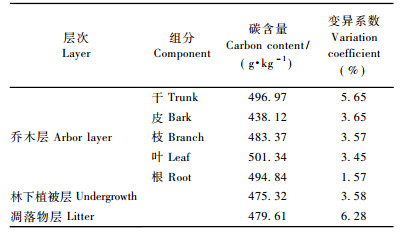
由表 3可见:土壤碳含量相对较小(表 3),各土层碳含量3.11~6.33 g·kg-1,表层(0~20 cm)土壤碳含量是亚表层(20~40 cm)土壤碳含量的1.35倍,是深层(80~100 cm)的2.03倍,土壤碳含量随着土壤深度的增加逐渐减小;不同研究地点之间各土层碳含量变异较大,随土壤深度加深变异逐渐减小。
|
|
表 4为海南岛不同区域各土层碳含量变异性,从表层(0~20 cm)土壤碳含量可见,中部山地土壤碳含量最高,达24.50 g·kg-1,其次为北部平原区,平均达10.80 g·kg-1,东部沿海台地区的土壤碳含量最小,仅为2.54 g·kg-1;从表层土壤碳含量的变异程度来看,中部山地变异系数最小,其次为北部平原区,西部沿海台地的变异系数最大。从不同区域各土层碳含量的变化来看,中部山地区和北部平原区的土壤碳含量均表现为随着土壤深度的增加而减小,但东部和西部沿海台地区的各土层碳含量变化不明显,其碳含量差异不大。
|
|

根据测定结果,海南尾细桉人工林调查样地乔木层总生物量平均为42.31 t·hm-2,其中树干生物量最大,达21.84 t·hm-2,占乔木总生物量的51.61%,树叶生物量次之,占总乔木层生物量的16.68%,树皮、树枝和树根3者的生物量相当,3者共占总乔木层生物量的31.7%(表 5)。
|
|
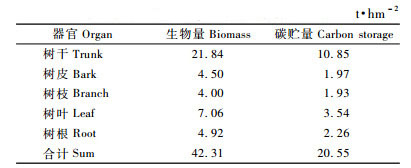
尾细桉各器官的生物量与相应碳含量的积为各器官碳贮量。由表 5可见,尾细桉人工林乔木层碳贮量平均为20.55 t·hm-2,其中树干碳贮量平均为10.85 t·hm-2,占乔木层碳贮量的52.81%,其次为树叶碳贮量,占乔木层碳贮量的17.22%,树皮、树枝和树根的碳贮量占乔木层碳贮量的29.96%。乔木层各组分碳贮量排序为树干(52.81%)>树叶(17.22%)>树根(10.98%)>树皮(9.58%)>树枝(9.40%)。
3.3 尾细桉人工林生态系统碳贮量表 6为尾细桉人工林生态系统各组分的生物量和碳贮量,结果表明,尾细桉人工林生态系统碳贮量平均为88.84 t·hm-2,乔木层碳贮量为20.55 t·hm-2,占23.13%;林下植被层碳贮量为1.55 t·hm-2,占1.74%;凋落物层碳贮量为1.93 t·hm-2,占2.17%;土壤(0~100 cm)碳贮量最大,为64.81 t·hm-2,占72.96%。土壤是尾细桉人工林生态系统的最大碳库,不同土层碳库具有一定的差异,表层(0~20 cm)土壤的碳贮量达19.94 t·hm-2,与乔木层碳贮量相当,亚表层(20~40 cm)土壤次之,达15.87 t·hm-2,80~100 cm土层碳贮量最小,仅为8.75 t·hm-2。
|
|

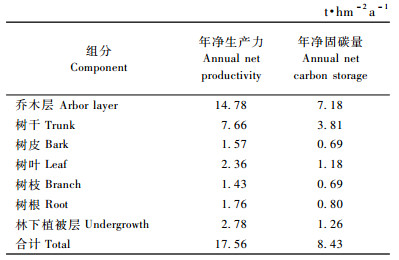
由表 7可见,海南尾细桉林生态系统的净生产力平均为17.56 t·hm-2a-1,年净固碳量平均为8.43 t·hm-2a-1,其中乔木层年固碳量为7.18 t·hm-2a-1,占85.10%。在乔木层中,以树干年固碳量最高,达3.81 t·hm-2a-1,占45.15%;其次为树叶,占14.04%,树皮最小,占8.15%。林下植被固碳量为1.26 t·hm-2a-1,占14.90%。
|
|
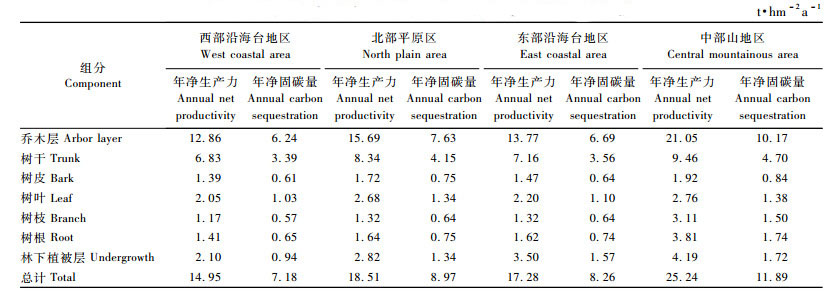
研究表明(表 8),海南中部山地区尾细桉人工林乔木层年生产力最大,达21.05 t·hm-2a-1,年固碳量达10.17 t·hm-2a-1,西部地区尾细桉人工林年生产力较低,仅为12.86 t·hm-2a-1,年固碳量为6.24 t·hm-2a-1。平均数差异性t检验结果表明,中部山地区尾细桉人工林生产力与西部沿海台地区、北部平原区和东部沿海台地区尾细桉林的生产力差异显著(P值分别为0.007 7, 0.043 6和0.006 6,P < 0.01),西部台海台地区、北部平原区和东部沿海台地区尾细桉林之间的生产力差异不显著(P>0.05)。林下植被年固碳能力以中部山地区最大,达1.72 t·hm-2a-1,西部沿海台地区最小,仅为0.94 t·hm-2a-1。植被层总固碳能力以中部山地区最大(11.89 t·hm-2a-1),西部沿海台地区最低(7.18 t·hm-2a-1)。平均数差异性t检验结果表明,中部山地区尾细桉人工林总固碳能力与西部沿海台地区、北部平原区和东部沿海台地区的总固碳能力差异显著(P值分别为0.000 25, 0.005 025和0.010 89,P < 0.05),西部沿海区、北部平原区和东部沿海台地区之间的总固碳能力差异不显著(P>0.05)。
|
|
由表 9可见:尾细桉人工生态系统碳贮量以中部山地区最大,达179.94 t·hm-2,东部沿海台地区最小,仅为55.26 t·hm-2。不同区域尾细桉林生态系统碳贮量差异性t检验结果表明,除东部沿海台地区和西部沿海台地区碳贮量差异不显著外,其他区域之间总碳贮量均差异显著(P<0.01)。
|
|
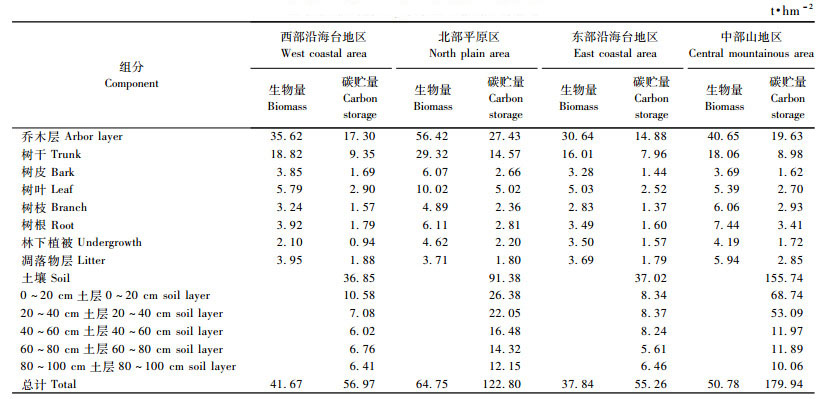
乔木层碳贮量以北部平原区最高,达27.43 t·hm-2,东部沿海台地区最小,仅为14.88 t·hm-2(表 9)。平均值差异显著性t检验(表 9)表明,北部平原区乔木层碳贮量与西部沿海台地区、东部沿海台地区、中部山地区之间差异较显著(P<0.1)。乔木层碳贮量主要与尾细桉人工林林龄及其生产力水平有关,北部平原区尾细桉人工林种植时间较长,林龄较大,中部山地区尾细桉人工林林龄较小,其生物量相对较低,而西部沿海台地区较为干旱,东部沿海台地区尾细桉主要种植于沙地上,其生产力均较低,所以尾细桉人工林乔木层碳贮量以北部平原区最大。不同区域尾细桉人工林林下植被层碳贮量以北部平原区最大,西部沿海台地区最小。凋落物碳贮量以中部山地区最大,达2.85 t·hm-2,北部平原区、东部沿海台地区和西部沿海台地区基本相当,均在1.80 t·hm-2左右(表 9)。
各区域间土壤碳贮量差异较大,中部山地区尾细桉人工林土壤碳贮量最大,达155.74 t·hm-2,北部平原区土壤碳贮量次之,为91.38 t·hm-2,东部沿海台地和西部沿海台地区土壤碳贮量相当,仅为33.0 t·hm-2左右(表 9)。土壤碳贮量平均值差异性t检验结果表明,除西部沿海台地区和东部沿海台地区之间不显著(P > 0.05)外,其他区域之间土壤碳贮量差异均极显著(P < 0.01)。中部山地区尾细桉人工林是在原天然林破坏后种植的,原土壤碳密度较大,北部平原区尾细桉人工林主要为灌丛地破坏后种植的尾细桉林,其土壤条件也较好,碳含量也相对较高,西部沿海台地区由于土壤水分含量低,且多为沙地,东部沿海台地区也为沙地,土壤碳积累量小,其碳密度很小。
4 结论与讨论海南尾细桉人工林生态系统碳贮量平均为88.84 t·hm-2,远低于尖峰岭热带山地雨林(340.467 t·hm-2)和天然更新林(258.996 t·hm-2)生态系统碳贮量(吴仲民等,1998)。整个海南尾细桉人工林(33.3万hm2)生态系统碳贮量为2 958.37万t C,其中乔木层为684.32万t C,林下植被层为51.61万t C,凋落物层为64.27万t C,土壤层为2 158.17万t C。
海南尾细桉人工林植被层碳贮量平均为24.03 t·hm-2,乔木层占植被层总碳贮量的85.52%,与广西东门林场2代林3.6年生尾细桉林植被层碳贮量相当(20.42 t·hm-2)(梁宏温等,2009),远低于我国森林植被平均碳贮量(57.07 t·hm-2)(周玉荣等,2000)及尖峰岭山地原始雨林植被碳贮量(232.791 t·hm-2)和自然更新林植被碳贮量(150.203 t·hm-2)(吴仲民等,1998)。本研究植被碳贮量较低的原因主要是尾细桉人工林林龄较低,平均仅为2.8年。
海南尾细桉人工林土壤(0~100 cm)有机碳贮量平均为64.81 t·hm-2,占整个生态系统碳贮量的72.96%,是植被碳贮量的2.70倍,远低于海南尖峰岭山地雨林和季雨林土壤(0~100 cm)碳贮量(97.10~119.54 t·hm-2,平均为102.60 t·hm-2) (吴仲民等,1997)。土壤是一个十分重要的碳库,我国森林土壤平均碳贮量为193.55 t·hm-2,约为植被碳贮量的3.4倍(周玉荣等,2000)。本研究中海南尾细桉人工林土壤碳贮量及其与植被碳贮量比值均低于全国平均水平,主要原因为:海南主要地处热带地区,水热条件相对较好,其生产力较高,有利于植被生物量积累,而土壤呼吸速率较高,凋落物分解速率较快,加之植被对土壤养分的大量吸收,造成土壤碳贮量相对较低(周玉荣等,2000);海南尾细桉人工林主要分布于原农业用地和土壤相对较贫瘠的土地(如沙地、旱地),土壤受人为干扰导致碳贮量较低或本身碳贮量较低;造林时炼山清除前茬采伐剩余物和林下植被,也导致土壤碳大量流失,这些均是海南尾细桉人工林平均土壤碳贮量较低的原因。
海南尾细桉人工林净生产力平均为17.56 t·hm-2a-1,年净固碳量平均为8.43 t·hm-2a-1,折算成CO2量为30.91 t·hm-2a-1,远高于尖峰岭热带山地雨林(3.817 t ·hm-2a-1)(李意德等,1998a)、11年生杉木(Cunninghamia lanceolata)林(3.489 t·hm-2a-1)(方晰等,2002)、苏南地区27年生杉木林(2.36 t·hm-2a-1)(阮宏华等,1997)、重庆46年生马尾松(Pinus massoniana)林(4.49 t·hm-2a-1)(张治军等,2009)、18年生樟树(Cinnamomum camphora)人工林(4.98 t·hm-2a-1)(雷丕锋等,2004)、19年生湿地松(Pinus elliottii)人工林(4.54 t·hm-2a-1)(涂洁等,2007)和32年生楠木(Phoebe bourmei)林(4.25 t ·hm-2a-1)(马明东等,2007)的固碳量,说明尾细桉人工林的固碳能力强,是较好的碳汇林业树种。整个海南尾细桉人工林(33.3×104 hm2)年固碳量达280.97×104 t C,相当于固定1 030.23×104 t CO2。
海南不同区域尾细桉人工林固碳能力存在一定的差异性,其中以中部山地区尾细桉人工林固碳能力最强,远远高于北部平原、西部沿海台地和东部沿海台地区,而西部沿海台地区、北部平原区和东部沿海台地区的尾细桉林固碳能力差异不大。主要原因为:中部山地区降雨量大,土壤多为壤土,水肥条件好,有利于尾细桉快速生长;北部平原区和东部沿海台地区降雨量也较大,但北部地区尾细桉人工林多种植于原农业用地,东部地区多为沿海沙地,土壤养分条件相较于北部平原地区差;海南西部沿海台地区处于季风的背阴面,降水量较小,蒸发量大,土壤干旱比较严重,尾细桉生长受制于水分条件,故其生产力最低。
不同区域尾细桉人工林碳贮量差异较明显,主要是由于土壤的碳贮量空间差异引起的,从显著性检验结果来看,西部沿海台地区和东部沿海台地区之间土壤碳贮量相当,其他区域之间土壤碳贮量差异较大。中部山地区尾细桉人工林前茬多为天然林或天然灌丛,土壤中原贮存的碳保存量较高,而北部平原区的前茬多为尾细桉林或灌丛,西部沿海台地区和东部沿海台地区的前茬多为甘蔗(Saccharum officinarum)或桉树(Eucalyptus spp.)林,土壤类型多为沙质或砂质土壤,土壤有机质保存较困难,故中部山地尾细桉人工林碳贮量最高,东部沿海台地区和西部沿海台地区较低。
对比本研究与尖峰岭热带雨林区(吴仲民等,1998;李意德等,1998b)结果表明,海南中部山地区是非常重要的高固碳能力与碳贮量区域,特别是土壤,中部山地区尾细桉人工林多为热带天然林或次生林破坏后营造起来的,但尾细桉人工林的碳贮量却远低于热带雨林和其自然更新林,因此海南中部区域应该严格加强天然林保护,以减少热带雨林生态系统碳释放,而海南北部平原、西部沿海台地及东部沿海台地区可以发展尾细桉人工林。
茶正早, 黎仕聪, 林钊沐, 等. 1999. 海南岛桉林土壤肥力的研究[J]. 热带作物学报, 20(2): 37-43. |
陈秋波. 2001. 尾细桉人工林生物多样性研究进展[J]. 热带作物学报, 22(4): 82-90. |
方晰, 田大伦, 项文化. 2002. 速生阶段杉木人工林碳素密度、贮量和分布[J]. 林业科学, 38(3): 14-20. DOI:10.11707/j.1001-7488.20020303 |
雷丕锋, 项文化, 田大伦, 等. 2004. 樟树人工林生态系统碳贮量与分布研究[J]. 生态学杂志, 23(4): 25-30. |
李意德, 吴仲民, 曾庆波, 等. 1998a. 尖峰岭热带山地雨林群落生产和二氧化碳同化净增量的初步研究[J]. 植物生态学报, 22(2): 127-134. |
李意德, 吴仲民, 曾庆波, 等. 1998b. 尖峰岭热带山地雨林生态系统碳平衡的初步研究[J]. 生态学报, 18(4): 371-378. |
梁宏温, 温远光, 温琳华, 等. 2009. 连栽对尾巨桉短周期人工林碳贮量的影响[J]. 生态学报, 29(8): 4242-4250. |
马明东, 江洪, 罗承德, 等. 2007. 四川西北部亚高山云杉天然林生态系统碳密度、净生产量和碳贮量的初步研究[J]. 植物生态学报, 31(2): 305-312. DOI:10.17521/cjpe.2007.0035 |
阮宏华, 姜志林, 高苏铭. 1997. 苏南丘陵主要森林类型碳循环研究—含量与分布规律[J]. 生态学杂志, 16(6): 17-21. |
唐红侠, 韩丹, 赵由才, 等. 2009. 农林业温室气体减排与控制[M]. 北京: 化学工业出版社.
|
涂洁, 刘琪璟. 2009. 亚热带红壤丘陵区湿地松人工林生态系统碳素贮量与分布研究[J]. 江西农业大学学报, 29(1): 48-54. |
吴仲民, 曾庆波, 李意德, 等. 1997. 尖峰岭热带森林土壤C储量和CO2排放量的初步研究[J]. 植物生态学报, 21(5): 416-423. |
吴仲民, 李意德, 曾庆波, 等. 1998. 尖峰岭热带山地雨林C素库及皆伐影响的初步研究[J]. 应用生态学报, 9(4): 341-344. |
张治军, 张小全, 王彦辉, 等. 2009. 重庆铁山坪马尾松林生态系统碳贮量及其分配特征[J]. 林业科学, 45(5): 49-53. DOI:10.11707/j.1001-7488.20090507 |
中国工程院桉树项目组. 2009. 科学种植尾细桉与环境保护(一). [EB/OL]. [2010-10-05]. http: //www.gdly.com/mk_direct.php?id =4926.
|
中国农业化学专业委员会. 1983. 农业土壤化学分析方法[M]. 北京: 科学出版社: 272-274.
|
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522. |
Marland E, Maland G. 2003. The treatment of long-lived, carbon-containing products in inventories of carbon dioxide emissions to the atmosphere[J]. Environmental Science & Policy, 6(2): 139-152. |
West T O, Marland G. 2002. A synthesis of carbon sequestration, carbon emissions and net carbon flux in agriculture: comparing tillage practices in the United States[J]. Agriculture, Ecosystems & Environment, 91(1/3): 217-232. |