 2011, Vol. 47
2011, Vol. 47文章信息
- 安玉艳, 梁宗锁, 韩蕊莲
- An Yuyan, Liang Zongsuo, Han Ruilian
- 黄土高原3种乡土灌木的水分利用与抗旱适应性
- Water Use Characteristics and Drought Adaptation of Three Native Shrubs in the Loess Plateau
- 林业科学, 2011, 47(10): 8-15.
- Scientia Silvae Sinicae, 2011, 47(10): 8-15.
-
文章历史
- 收稿日期:2010-06-18
- 修回日期:2011-09-05
-
作者相关文章



2. 中国科学院水土保持与生态环境研究中心 杨凌 712100
2. Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences Yangling 712100
黄土高原是我国典型的干旱半干旱地区,该区降水量集中且偏少,水土流失严重,植被覆盖情况差(Li et al., 2008),全球气候变化还将加剧黄土高原的水分亏缺程度(Coley, 1998; Wassmann et al., 2009)。造林种草是治理水土流失、改善生态环境的主要生物措施,而水分亏缺是制约该区植被恢复和治理的关键因素(张淑勇等, 2008; 张建军等, 2009)。黄土高原的乡土植物经过长期的自然选择和协同进化生长良好并能自行繁衍,这些乡土植物种无疑是营造人工林、加快植被恢复的最合适资源,但目前人们对这些乡土植物的生物学特性、耗水规律及抗旱适应性的认识还十分缺乏,限制了一些优良乡土树种资源的开发和利用。
杠柳(Periploca sepium)、丁香(Syringa oblata)和连翘(Forsythia suspense)是黄土丘陵区广泛分布的乡土灌木树种,均具有一定的观赏价值、药用价值和生态功能(任宪威, 1997),在黄土高原退耕还林与植被恢复中,具有较大的开发潜力。目前对杠柳的研究多集中在化学成分及经济价值上(王磊等, 2007; Bamba et al., 2007),对丁香和连翘的研究多集中在生物活性物质和城市园林绿化上(陈玉俊等, 1999; 张树军等, 2006; 尉小慧等, 2009),对3树种耗水规律及抗旱生理生化适应性的研究相对较少。本研究人工控制土壤水分,分析不同强度干旱胁迫对杠柳、丁香和连翘3树种的生长、耗水及生理生化特性的影响,比较3树种在干旱胁迫下的水分利用及抗旱适应性特征,揭示3树种适应黄土丘陵干旱半干旱环境的抗旱策略,为3树种在该区人工植被建设中的开发利用及科学合理配置提供理论依据。
1 材料与方法本试验选用黄土高原常见的乡土灌木杠柳、丁香和连翘的2年生天然实生幼苗(均采自陕西省安塞县中国科学院安塞水土保持试验站野生封育坡面),杠柳平均株高和地径分别为36.0和0.8 cm,丁香平均株高和地径分别为26.25和0.85 cm,连翘平均株高和地径分别为28.01和0.49 cm。
选择大小基本一致的苗木于2006-03植入高27 cm,上口径35 cm,下口径22 cm的塑料圆桶中,桶中均装过筛垆土,田间最大持水量(FC)为28%,桶底部铺有石头和滤纸,内插一塑料管,便于从底部浇水,避免土壤板结。盆栽桶放置于中国科学院水土保持研究所的可移动防雨棚下,雨天用防雨棚遮雨,晴天露地生长。并用塑料薄膜覆盖盆中裸土,同时设置裸土对照盆(不栽植物),以排除土壤蒸发,栽植苗木后正常浇水使之萌芽,生长2个月左右,进行水分控制,待土壤水分降至试验要求后,用电子秤(载量30 kg,感量5 g)称重法将土壤含水量控制在设定范围内,并准确记录加水量。试验设置3个水分处理:75%FC;55%FC;40%FC。每个水分处理均重复6次,每桶选留1株。整个试验持续至10月中旬结束,历时225天。
5月20日开始控制土壤水分进行水分处理。用电子秤每2天定时称盆质量,在排除土壤蒸发后,以盆质量的减少量为树种在该阶段的耗水量,并加水补充至设定土壤含水量。月耗水量为当月每日耗水量的总和(其中5和10月的月耗水量为当月的日均耗水量与当月天数31的乘积);总耗水量为各月耗水量之和。用电子秤每天定时称量并记录盆质量,排除苗、桶、干土、石头以及滤纸和塑料管质量后即为当时该盆的水分质量,该水分质量占干土质量百分比即为当时土壤含水量。本试验中,3个水分处理下杠柳的土壤水分含量分别为:20.4%±1.1%(75% FC),15.6%±1.0%(55% FC)和10.9%±0.9%(40% FC);丁香分别为20.7%±1.1%(75% FC),15.7%±0.6%(55% FC)和11.2%±0.7%(40% FC);连翘分别为20.2%±1.5%(75% FC),15.2%±1.1%(55% FC)和10.6%±0.9%(40% FC)。方差分析表明在同种水分处理下3树种的土壤水分含量无显著差异,这是进行3树种生长、水分利用及抗旱性比较的前提。
新生枝条生长速率:5月1日开始水分处理后选取每株植物中上部的新生枝条挂牌标记,每个处理6个植株,每株选取5个枝条,每隔几天用毫米刻度尺于上午9:00测定幼苗新生枝条长度,测定频率根据枝条生长快慢进行调整;单叶叶面积:用CI-202手持式CCD扫描叶面积仪测量完全成熟叶片的面积,每个处理测量20个叶片,取平均值作为该处理单叶叶面积;试验结束时将苗木连根拔起,小心冲洗干净后分成根、茎、叶3部分分别烘干,用于计算根冠比;试验结束时苗木的总干质量减去栽前干质量即为生长季积累的生物量,栽前干质量的测定是选取与将要移栽的苗木大小一致的苗木10株,烘干称质量,取其平均值。水分利用效率(WUE)为生长季生物量增量与生长季耗水总量的比值。
植物体对水分亏缺的生理反应不仅与胁迫强度有关,还与胁迫时间以及植物体自身的生长发育阶段有密切关系(Bray, 1997),本研究中所有生理指标于试验处理40天时开始测定,每隔20天重复测定1次,共测定6次(7月1日,7月20日,8月10日,8月30日,9月20日和10月10日),每次测定均做3次重复。测定各生理指标时每盆至少选取2片叶子。
叶片相对含水量(Crw)的测定:取最新完全展开的叶片,测定叶片鲜质量(Fw)、叶片置于4 ℃完全黑暗的环境中吸水6 h后的饱和质量(Tw)及叶片置于60 ℃烘箱中48 h后的质量(Dw),Crw (%) =(Fw-Dw)× 100/(Tw-Dw)。叶片保水力的测定:自然干燥法测定,取功能叶在离体条件下饱和吸水6 h后,自然缓慢脱水24 h,称质量(W1),将叶片在80 ℃下烘干,称取干质量(Wd),以自然缓慢脱水24 h时的叶片含水量(占干质量)衡量保水力。整个过程均在28 ℃恒温干燥培养箱中进行。
酶液提取方法:称取植株中上部最新完全展开的杠柳叶片鲜样0.5 g于预冷研钵中,加入2 mL预冷的提取介质(内含0.1% PVP的50 mmol·L-1 pH 7.8 477201220的磷酸缓冲液)及少量石英砂在冰浴下研磨成匀浆,加入提取介质冲洗研钵2~3次,并使最终体积为10 mL,于4 ℃下10 000 r·min-1离心15 min,上清液即为酶提取液,于冰箱中保存备用,该酶提取液用于SOD,POD,MDA及可溶性蛋白质含量测定。SOD活性用氮蓝四唑光还原法测定;POD活性用愈创木酚显色法测定;MDA含量用硫代巴比妥酸比色法测定;细胞质膜透性采用电导仪法测定(高俊凤, 2000)。游离脯氨酸含量用磺基水杨酸法提取茚三酮比色法测定(Bates, 1973);可溶性糖含量用蒽酮比色法测定(Yemm et al., 1954);可溶性蛋白含量用考马斯亮蓝G-250染色法测定(Bradford, 1976)。
所有试验处理的生长、生理数据利用SPSS(16.0)进行单因素方差分析(ANOVA),对处理之间以及同一处理随时间变化的差异性多重比较采用LSD (least significant difference)分析,分别设定α=0.05和α=0.01为显著性和极显著性水平。所有图片均在Origin(7.0)或Excel(2003)软件系统下完成。
2 结果与分析 2.1 苗木耗水规律由图 1可以看出,3树种的月耗水量均表现为75%FC>55%FC>40%FC,方差分析表明3种不同强度干旱胁迫下3树种的耗水量差异达极显著水平。3树种耗水量在3种土壤水分条件下均表现为连翘>杠柳>丁香。整个生长季3个树种在3种水分条件下的月耗水变化均呈单峰曲线,3树种在6—8这3个月的耗水量均达整个生育期总耗水量的70%以上。

|
图 1 3树种的单株月耗水量 Figure 1 Monthly water consumption of single plant |
不同强度干旱胁迫下3树种生长速率均表现为75%FC>55%FC>40%FC(图 2),且不同处理间差异显著。不同树种新生枝条生长速率差异显著,连翘>杠柳>丁香。生长速率的差异使最终的枝条长差异显著(表 1)。如75%FC条件下,连翘的枝条长约为杠柳的1.6倍、丁香的2.4倍。

|
图 2 新生枝条伸长生长动态 Figure 2 Changes of the new branch's length |
|
|
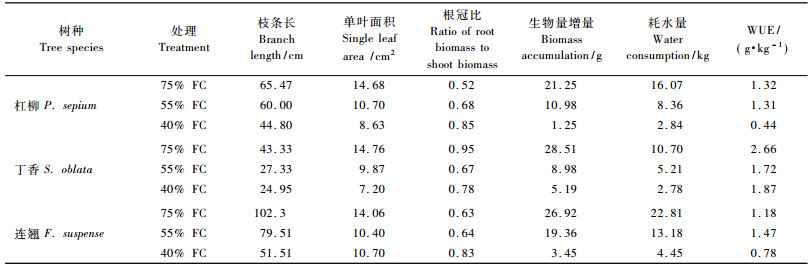
杠柳和丁香的叶面积均随干旱强度的加剧显著减小(表 1),丁香的降低幅度更大,表明其对水分胁迫的反应更敏感。连翘叶面积从75%FC到55%FC显著性降低,但从55%FC到40%FC无明显变化。3树种在整个生长季的生物量增量表现一致,均随土壤水分含量的减少显著降低。
除丁香在75%FC下的根冠比超过55%FC和40%FC条件下外,3树种其余各处理根冠比均随干旱强度加剧逐渐增大(表 1)。杠柳根冠比增大幅度最大,与75%FC相比,在55%FC和40%FC土壤水分条件下杠柳的根冠比分别增加了30.8%和63.5%。连翘的根冠比只在土壤水分严重亏缺(40%FC)时显著增大。
2.3 苗木水分利用效率土壤水分含量对不同树种水分利用效率的影响不同(表 1)。丁香的水分利用效率在75%FC下最高,干旱胁迫加剧至55%FC时极显著降低,从55%FC到40%FC其水分利用效率无显著变化。杠柳在75%FC和55%FC下的水分利用效率无明显差异,在40%FC下的水分利用效率极显著下降。连翘在55%FC条件下的水分利用效率最高,75%FC下次之,40%FC下最低,且3种土壤水分下的WUE差异显著。
2.4 苗木叶片生理指标丁香的叶片含水量(表 2)在3种水分条件下无明显差异,但与杠柳和连翘的叶片含水量差异显著;同种水分条件下杠柳的叶片含水量显著低于连翘。3树种叶片相对含水量受土壤水分含量影响显著(表 2)。与75%FC相比,55%FC和40%FC水分条件下杠柳叶片相对含水量在整个生长季的平均值分别下降了9.89%和22.55%;丁香分别下降了12.50%和21.81%;连翘分别下降了10.79%和19.57%。3树种叶片保水力(表 2)对不同强度干旱胁迫响应不同。杠柳叶片保水力随干旱胁迫加剧而显著性增强;丁香叶片保水力随干旱胁迫加剧则逐渐减弱,在40%FC条件下显著降低;连翘叶片保水力在55%FC下最高,显著高于75%FC和40%FC。3树种相比,杠柳叶片保水力远远大于丁香和连翘。
|
|
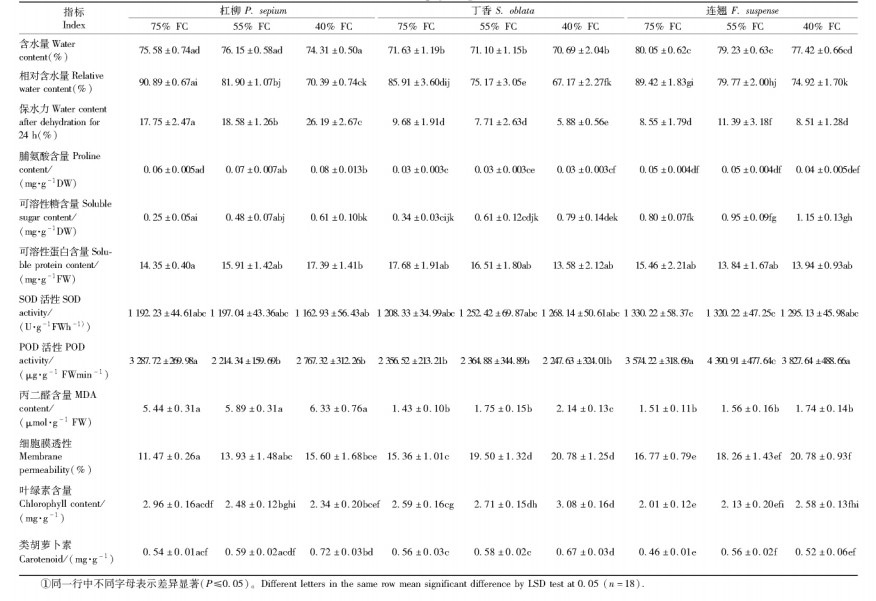
本试验用MDA含量和细胞膜相对透性来衡量脂质过氧化程度(表 2)。杠柳和连翘的MDA含量随干旱胁迫强度的增强逐渐升高(P>0.05);丁香的MDA含量只在土壤干旱至40%FC时显著升高。3树种相比,杠柳的MDA含量显著高于丁香和连翘。与MDA含量相反,在同种土壤水分条件下杠柳的细胞膜透性显著低于丁香,连翘的膜透性最大。从变化趋势上看,3树种的细胞膜透性均随土壤干旱胁迫的加剧而逐渐增大。
SOD和POD为植物体内2种重要的抗氧化保护酶。由表 2可以看出,3树种的SOD以及丁香的POD活性在3种水分条件下无显著差异。杠柳的POD活性在75% FC下最高,在55% FC和40% FC下显著降低,连翘在55% FC下的POD活性显著高于75% FC和40% FC。3树种的POD活性整体表现为连翘>杠柳>丁香。类胡萝卜素是植物体抵抗氧化胁迫的一种重要抗氧化物质。3树种在40%FC下的类胡萝卜素含量均显著高于75%FC。
由表 2脯氨酸含量可以看出,丁香和连翘的脯氨酸含量在3种水分条件下无明显差异,杠柳在40% FC下的脯氨酸含量显著高于75%FC下的含量。3树种相比,杠柳的脯氨酸含量最高,连翘次之,丁香最小。可溶性糖的含量则表现为连翘>丁香>杠柳。3树种的可溶性糖含量均表现为随干旱胁迫强度的增加而升高。杠柳在55% FC和40% FC下的可溶性糖含量分别为75%FC的1.92和2.44倍;丁香在55%FC和40%FC下的可溶性糖含量分别为75%FC的1.79和2.32倍;连翘在55%FC和40%FC下的可溶性糖含量分别为75%FC的1.19和1.44倍。3树种的可溶性蛋白含量在不同强度干旱胁迫下的变化不同。杠柳的可溶性蛋白含量随土壤水分的减少逐渐增加,丁香和连翘则相反。方差分析表明,杠柳在40% FC下的可溶性蛋白含量显著高于75% FC(P≤0.05),丁香和连翘的可溶性蛋白含量在3种土壤条件下的差异不显著。分析表 2中叶绿素含量可知,杠柳的叶绿素含量随水分亏缺加剧逐渐减少,丁香和连翘表现出相反的变化趋势。3树种在75%FC和40%FC下叶绿素含量的差异均达到显著性水平(P≤0.05)。
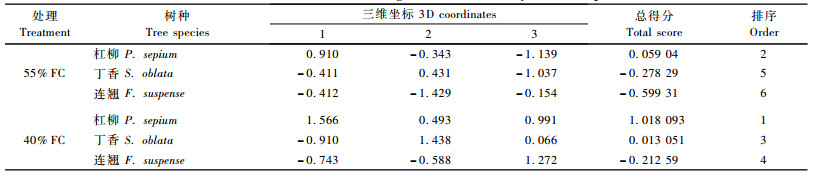
2.5 苗木抗旱适应性综合评价从各生理指标的试验结果可以看出,不同的生理生化指标在反映同一植物的抗逆性表现时可能会有所不同,且植物对干旱胁迫的响应与干旱胁迫强度密切相关。本研究选择了11个指标:叶片相对含水量、叶片保水力、叶片脯氨酸含量、叶片可溶性糖含量、叶片可溶性蛋白含量、叶片POD活性、叶片细胞膜透性、叶片叶绿素含量、叶片类胡萝卜素含量、根冠比和水分利用效率,采用主成分分析方法对55%FC和40%FC这2种干旱胁迫下的杠柳、丁香和连翘苗木的抗旱性进行了综合评价。
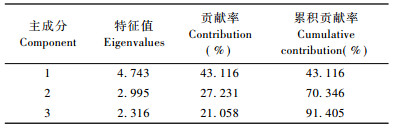
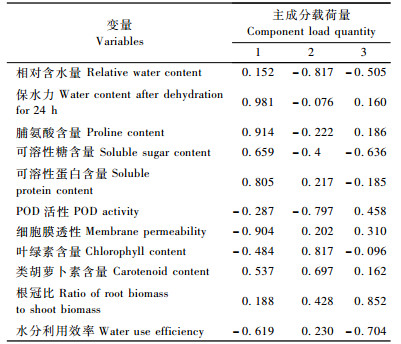
主成分分析是将多个指标划归为少数几个相互独立指标的一种统计方法,其在林木抗旱性研究中,尤其在确定影响林木抗旱性的主要指标的研究中被广泛应用(朱春云等, 1996; 王青宁等, 2005)。由表 3可见,本研究共提取了3个主成分,其累积贡献率达91.41%,说明这3个综合指标能够很好地代表原来11个指标的特性。由表 4可以看出,第1主成分上较大的特征向量分为2组,一组是保水力、脯氨酸含量、可溶性蛋白含量和可溶性糖含量,后3者均为渗透调节物质,有助于降低胞内渗透势,提高吸水力,保水力与吸水力都反映了植物的御旱特性;另一组是细胞膜透性,但其属于“下降指标”,反映的是干旱下植物稳定细胞膜的能力,进一步反映了植物的耐旱特性,因此可称第1主成分为抗旱因子。第2主成分上载荷较高且符号为正的指标有叶绿素含量和类胡萝卜素含量,载荷较高且符号为负的指标有叶片相对含水量和POD活性,这些指标反映了植物在体内水分减少时抵抗氧化胁迫的能力,故称第2主成分为抗氧化因子。第3主成分主要受根冠比和水分利用效率支配,2者反映了生物量积累与分配特征,故称这一主成分为生物量因子。
|
|
|
|
由表 5可以看出,3树种的抗旱性大小顺序为:杠柳>丁香>连翘。3树种在40%FC下的总得分均高于55%FC,说明3树种在土壤水分从55%FC降到40%FC时抗旱适应性更强了,表明3树种都具有较强的抗旱能力。
|
|
3种水分条件下3树种的耗水高峰均出现在6—8这3个月(图 1),而7月和8月阶段性雨热同步是陕北黄土丘陵区典型的气候特征(杨新国等, 2008),6—8这3个月也正是3树种的快速生长期,可见,3树种的耗水高峰期、快速生长期与其生长环境的雨热时期是重叠的。杨建伟等(2004a; 2004b; 2006)对黄土高原常用造林外来种的研究表明:杨树(Populus spp.)和刺槐(Robinia pseudoacacia)的快速生长期正是黄土高原雨季来临前土壤水分最为亏缺的时期,这导致杨树林、刺槐林生长发育不良,形成大面积“小老树”,且杨树、刺槐等为高耗水树种,容易造成“土壤干层”的形成。与外来种比较,3种乡土灌木能够充分利用黄土丘陵区有限的雨热资源快速积累生物量,这可能是3树种在黄土丘陵区普遍分布的一个重要原因。
本研究表明,随干旱胁迫加剧,3树种叶面积减小,且茎和叶片生物量所占比重普遍降低,这一方面降低了蒸腾面积,减慢了植物体内水分的散失,一定程度上缓解了干旱胁迫;另一方面,降低了光合面积和生长潜力;在干旱胁迫下,随着土壤水分含量的减少,3树种均倾向于将更多的碳分配到根部,壮大根系,以提高根系对矿物质和水分的吸收(Edwards et al., 2004),以杠柳表现最为突出。
不同的指标对树种的抗旱性贡献不同(谢建坤等, 2010),故在评价树种抗旱性的过程中,不能单独依靠上述某个或某2个指标(Ruiz-Sanchez et al., 2000; Prasil et al., 2007),必须根据一系列生理生化指标的重复测定和综合评定才能提高鉴定结果的准确性(王宇超等, 2010),从整体上揭示植物的抗性能力。
本研究中,主成分分析从11个生理指标中提取了3个主成分,即抗旱因子、抗氧化因子和生物量因子。3树种在3个主成分上的得分值不同,杠柳在抗旱因子上的得分远大于丁香和连翘,在抗氧化因子上得分最高的是丁香,其次为杠柳,而在生物量因子上得分最高的则是连翘,其次为杠柳。由这些结果分析得知,3树种抗旱策略各有不同。杠柳叶片保水力强,水分散失缓慢,且干旱胁迫下主动积累脯氨酸、可溶性糖和可溶性蛋白等有机渗透调节物质,降低渗透势,提高吸水力,从而能够防御或者延缓水分胁迫给植物体造成的伤害。另外,杠柳的细胞膜透性在3树种中最低,主成分分析表明,杠柳的细胞膜对干旱胁迫具有较强的耐受性,这是杠柳适应干旱环境的又一重要生理机制。干旱条件下杠柳脯氨酸等溶质的积累均有助于细胞膜的稳定(Verbruggen, et al., 2008)。第2主成分主要受叶绿素和类胡萝卜素含量正向支配,丁香在此主成分上得分最高。李清芳等(2007)研究表明,干旱胁迫会抑制叶绿素的生物合成、提高叶绿素酶活性并加速叶绿素分解,导致叶绿素含量显著下降。本研究中,在干旱条件下,丁香叶片叶绿素以及类胡萝卜素含量均呈升高趋势(表 2)。相似的结果也出现在轻度干旱胁迫下的沙地云杉(Picea mongolica)中(邹春静等, 2003)。类胡萝卜素是植物体内一种重要的抗氧化剂,干旱条件下丁香叶片内类胡萝卜素含量的升高有助于缓解氧胁迫对其光系统的伤害,同时类胡萝卜素还参与光反应中心中过剩光能的耗散(邓馨等, 2000),保护光合机构。因此较强的光合抗氧化能力是丁香适应干旱环境的重要机制。连翘在严重干旱条件下根冠比显著增大,主成分分析显示连翘的抗旱性与其根冠比在干旱下的增大密切相关。这表明连翘在干旱下主要依赖于壮大根系,增强水分吸收以抵抗干旱胁迫。在40%FC条件下连翘水分利用效率的降低与其叶片比重的降低即光合面积的减小密切相关。结合连翘的生长特点发现,连翘在土壤水分>55%FC时生长迅速,固碳能力强,但当土壤水分继续干旱时,连翘则牺牲生物学产量和水分利用效率,提高其在干旱下的生存几率。这应该是连翘对黄土丘陵干旱半干旱环境的适应和生存策略。
陈玉俊, 项进, 许美娟. 1999. 连翘化学成分的研究[J]. 中国中药杂志, 24(5): 296-297. |
邓馨, 胡志昂, 王洪新, 等. 2000. 脱水和复水对复苏植物牛耳草离体叶片光合作用的影响[J]. 植物学报, 42(3): 321-323. |
高俊凤. 2000. 植物生理学实验技术[M]. 西安: 世界图书出版公司.
|
李清芳, 马成仓, 尚启亮. 2007. 干旱胁迫下硅对玉米光合作用和保护酶的影响[J]. 应用生态学报, 18(3): 531-536. |
任宪威. 1997. 树木学[M]. 北京: 中国林业出版社.
|
王磊, 殷志琦, 张雷红, 等. 2007. 杠柳根皮化学成分研究[J]. 中国中药杂志, 32(13): 1300-1302. DOI:10.3321/j.issn:1001-5302.2007.13.012 |
王青宁, 唐静, 衣学慧, 等. 2005. 基于多元统计评价毛白杨无性系的抗旱性[J]. 西北林学院学报, 20(4): 21-26. |
王宇超, 王得祥, 彭少兵, 等. 2010. 干旱胁迫对木本滨藜生理特性的影响[J]. 林业科学, 46(1): 61-67. DOI:10.11707/j.1001-7488.20100110 |
尉小慧, 张淑霞, 翟卫峰, 等. 2009. HPLC测定紫丁香树枝中橄榄苦苷的含量[J]. 中国中药杂志, 34(3): 304-306. |
谢建坤, 胡标林, 万勇, 等. 2010. 东乡普通野生稻与栽培稻苗期抗旱性的比较[J]. 生态学报, 30(6): 1665-1674. |
杨建伟, 梁宗锁, 韩蕊莲, 等. 2004a. 不同土壤水分状况对刺槐的生长及水分利用特征的影响[J]. 林业科学, 40(5): 93-98. |
杨建伟, 梁宗锁, 韩蕊莲, 等. 2004b. 不同干旱土壤条件下杨树的耗水规律及水分利用效率研究[J]. 植物生态学报, 28(5): 630-636. |
杨建伟, 梁宗锁, 韩蕊莲, 等. 2006. 黄土高原常用造林树种水分利用特征[J]. 生态学报, 26(2): 559-566. |
杨新国, 李玉英, 吴天龙, 等. 2008. 半干旱黄土丘陵沟壑区柳枝稷(Panicum virgatum)的生物质形成[J]. 生态学报, 28(12): 6043-6050. DOI:10.3321/j.issn:1000-0933.2008.12.032 |
张建军, 张岩, 张波. 2009. 晋西黄土区水土保持林地的土壤水分[J]. 林业科学, 45(11): 63-69. DOI:10.11707/j.1001-7488.20091111 |
张树军, 张军锋, 王金兰. 2006. 紫丁香树皮的化学成分研究[J]. 中草药, 37(11): 1624-1626. DOI:10.3321/j.issn:0253-2670.2006.11.008 |
张淑勇, 周泽福, 张光灿, 等. 2008. 半干旱黄土丘陵区4种天然次生灌木光合生理和水分利用特征[J]. 林业科学, 44(12): 140-146. DOI:10.3321/j.issn:1001-7488.2008.12.025 |
朱春云, 赵越, 刘霞, 等. 1996. 锦鸡儿等旱生树种抗旱生理的研究[J]. 干旱区研究, 13(1): 59-63. |
邹春静, 韩士杰, 徐文铎, 等. 2003. 沙地云杉生态型对干旱胁迫的生理生态响应[J]. 应用生态学报, 14(9): 1446-1450. |
Bamba T, Sando T, Miyabashira A, et al. 2007. Periploca sepium Bunge as a model plant for rubber biosynthesis study[J]. Zeitschrift Fur Naturforschung C-a Journal of Biosciences, 62(7): 579-582. |
Bates L S. 1973. Rapid determination of free proline for water-stress studies[J]. Plant and Soil, 39(1): 205-207. DOI:10.1007/BF00018060 |
Bray E A. 1997. Plant responses to water deficit[J]. Trends in Plant Science, 2(2): 48-54. DOI:10.1016/S1360-1385(97)82562-9 |
Bradford M M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye bingding[J]. Analytical Biochemistry, 72: 248-254. DOI:10.1016/0003-2697(76)90527-3 |
Coley P D. 1998. Possible effects of climate change on plant/herbivore interactions in moist tropical forests[J]. Climatic Change, 39(2): 455-472. |
Edwards E J, Benham D G, Marland L A, et al. 2004. Root production is determined by radiation flux in a temperate grassland community[J]. Global Change Biology, 10(2): 209-227. DOI:10.1111/gcb.2004.10.issue-2 |
Li W, Wang Q J, Wei S P, et al. 2008. Soil desiccation for Loess soils on natural and regrown areas[J]. Forest Ecology and Management, 255(7): 2467-2477. DOI:10.1016/j.foreco.2008.01.006 |
Prasil I T, Prasilova P, Marik P. 2007. Comparative study of direct and indirect evaluations of frost tolerance in barley[J]. Field Crops Research, 102(1): 1-8. DOI:10.1016/j.fcr.2006.12.012 |
Ruiz-Sanchez M C, Domingo R, Torrecillas A, et al. 2000. Water stress preconditioning to improve drought resistance in young apricot plants. Plant Science, 156(2): 245-251.
|
Verbruggen N, Hermans C. 2008. Proline accumulation in plants: a review[J]. Amino Acids, 35(4): 753-759. DOI:10.1007/s00726-008-0061-6 |
Wassmann R, Jagadish S V K, Heuer S, et al. 2009. Climate change affecting rice production: the physiological and agronomic basis for possible adaptation strategies[J]. Advances in Agronomy, 101(8): 59-122. |
Yemm E W, Willis A J. 1954. The estimation of carbohydrates in plant extracts by anthrone[J]. Biochemistry, 57(3): 508-514. DOI:10.1042/bj0570508 |