 2011, Vol. 47
2011, Vol. 47文章信息
- 李庆, 江南, 杨群芳, 周建华, 肖银波, 虞超
- Li Qing, Jiang Nan, Yang Qunfang, Zhou Jianhua, Xiao Yinbo, Yu Chao
- 麻疯树柄细蛾交尾行为及雄虫性信息素粗提物的触角电位反应
- Mating Behavior and Male EAG Response to Crude Extracts of Sex Pheromones from Female Stomphastis thraustica
- 林业科学, 2011, 47(9): 88-92.
- Scientia Silvae Sinicae, 2011, 47(9): 88-92.
-
文章历史
- 收稿日期:2010-01-26
- 修回日期:2010-06-17
-
作者相关文章


2. 四川省林业科学研究院森林保护研究所 成都 610081
2. Institute of Forest Protection, Sichuan Academy of Forestry Chengdu 610081
麻疯树(Jatropha curcas)为大戟科性能优良的生物柴油原料树种,是国际上公认的重要能源植物,有很高的经济价值(王秀娟等,2008)。麻疯树柄细蛾(Stomphastis thraustica)属鳞翅目(Lepidoptera),细蛾科(Gracillariidae),主要分布于印度、马来西亚、中非共和国、刚果、加纳、马达加斯加、尼日利亚、南非和津巴布韦。该虫潜叶危害麻疯树、棉叶麻疯树(Jatropha gossypifolia)、蓖麻(Ricinus communis)、卵叶白绒草(Leucas mollissima)和茶等。麻疯树人工林均可受害,1~3年生的有虫株率为50.5%~95.6%,严重时树叶全部脱落,连年危害可造成树势衰弱,严重影响麻疯树的生长和发育(周建华等,2008)。对该虫的研究主要集中在分类鉴定和生物学特性方面(肖银波等,2008)。
昆虫性信息素在害虫的综合治理方面具有极大的应用潜力(高文财等,2005)。在鳞翅目细蛾科昆虫中,除金纹细蛾(Lithoxolletis ringoniella)以外,未见对其他昆虫性信息素的研究报道。观察昆虫交尾行为和节律能为提取其性信息素提供时间和行为上的依据。触角电位技术(electroantennography,EAG)能准确地了解昆虫性信息素产生和释放的动态节律。为此,笔者仔细观察了麻疯树柄细蛾成虫的求偶和交尾行为、交尾节律,并对其性信息素的提取、雄虫对性信息素粗提物的EAG反应进行了研究,旨在为利用性信息素防治麻疯树柄细蛾提供理论依据。
1 材料与方法 1.1 供试虫源四川省林业科学研究院森林保护研究所提供麻疯树柄细蛾虫蛹。以盆栽麻疯树幼苗作为寄主进行饲养并建立种群。在(25±1)℃、光周期16L: 8D的RXZ型智能人工气候箱(宁波江南仪器厂)内繁殖多代。将蛹装入长7 cm、直径1 cm的指形管中单管培养,管口用保鲜膜封口并用针刺许多小孔,待其羽化后,按日龄(成虫羽化后0~24 h被视为1日龄)分别将雌、雄成虫置于放有树苗的不同纱笼内,笼中以10%的蜂蜜水饲养成虫并作为试验虫源。
1.2 麻疯树柄细蛾交尾行为及其动态节律的观察 1.2.1 麻疯树柄细蛾两性识别、交尾行为的观察在1 000 mL的烧杯中放入1日龄的健康雌雄成虫10对,在烧杯内用脱脂棉球蘸10%的蜂蜜水作为补充营养,用带针刺小孔的保鲜膜封住杯口,保证烧杯里的空气充足。观察雌雄成虫间的识别、交尾行为。
1.2.2 麻疯树柄细蛾交尾动态节律的研究在成虫羽化后,将15对1日龄处女成蛾放于1 000 mL的烧杯中进行交尾。在烧杯内用脱脂棉球蘸10%的蜂蜜水作为补充营养,用带针刺小孔的保鲜膜封住杯口。每10 min记录1次交尾情况,记录在此时段开始交尾的对数,每对交尾开始的时间以及交尾持续的时间,连续观察5天,试验重复3次,最后将数据按每小时的交尾频率进行统计(王惠等,1997)。
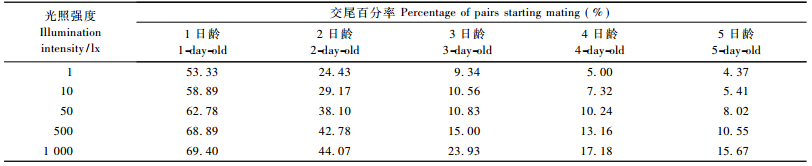
1.2.3 光照强度对麻疯树柄细蛾求偶交尾行为的影响在1 000 mL的烧杯中放入15对刚结束8 h黑暗处理的1日龄成虫,用带针刺小孔的保鲜膜封住杯口。光源用2 000 W白炽灯,在给光的一端,调整光源与放有成虫的烧杯之间的间距,使得供试成虫可获得不同强度的光照,并以照度计测定光照强度。光照强度被分别确定为1,10,50,500,1 000 lx(叶辉等,2006),每个处理重复3次,观察不同的光强信号对麻疯树柄细蛾求偶交尾的影响,连续观察5天。计算每个处理的交尾频率,统计每个处理中每对蛾子的交尾持续时间。
1.3 麻疯树柄细蛾雄蛾对性信息素粗提物的电生理反应(EAG) 1.3.1 雌性性信息素的腺体粗提参照陆鹏飞等(2007a),将10头1日龄有求偶行为的处女雌蛾腹部末端性腺剪下,置于100 μL的溶剂中,室温浸泡提取约1 h后,剔除管中腺体,并密封,放入-20 ℃冰箱保存。
1.3.2 雌性性信息素的空气收集参照陆鹏飞等(2007b),将1头1日龄有求偶行为的处女雌蛾放入一个指形管内,管口密封,静置20 min左右,向管中加入少量溶剂,充分摇匀,再迅速将溶液转移到另一洁净玻璃管中,密封保存于-20 ℃冰箱内。
1.3.3 性信息素粗提物提取适宜溶剂的确定采用腺体提取技术,提取性信息素的溶剂分别为2种弱极性的溶剂(正己烷、正戊烷),2种强极性的溶剂(二氯甲烷、乙醚)。以相应提取液作对照,分别用于EAG分析,检验提取效果,每个试验平行做3份。试验浓度为1 FE·(10 μL)-1,每次进样10 μL。
1.3.4 不同日龄雄蛾对1日龄处女雌蛾性信息素粗提物的EAG反应将10头1日龄有求偶行为的处女雌蛾用100 μL正己烷进行腺体提取。同时选取1头1日龄有求偶行为的处女雌蛾进行性信息素空气收集,提取液经N2进行浓缩。两者都以正己烷为对照进行触角电位反应测试。所用雄虫分别为活跃的1到4日龄,触角完好。试验浓度为1 FE·(10 μL)-1,每次进样10 μL。
1.3.5 性信息素粗提物的剂量反应曲线参照陆鹏飞等(2007)方法,将10头1日龄有求偶行为的处女雌蛾用100 μL正己烷进行腺体提取,溶液浓度为1 FE·(10 μL)-1。以此为母液分别配置1,10-1,10-2,10-3 FE·(10 μL)-1不同浓度的溶液,每处理进样10 μL,以正己烷为对照进行触角电位反应。
1.3.6 触角电位试验(EAG)将雄蛾触角切下,用导电胶将其连接在EAG探头中间; 用移液枪吸取一定量的粗提物,滴在2 cm×0.5 cm的滤纸上,然后将滤纸放入1 000 μL移液枪头内,枪头基部连接空气脉冲输出管口,端部插入连续空气输出管的小孔内,启动记录程序,混合了刺激物的气流吹到触角上,触角对其的反应被记录在电脑上。
触角电位仪(SYNTECH公司)包括微动操作台(SYNTECH MP-15)、刺激控制器(SYNTECH CS-55)、数据采集系统(SYNTECH IDAC-2)。试验时整个仪器置于金属屏蔽罩内。气流口离触角距离为0.5 cm,连续气体流量50 mL·s-1,刺激气体流量12.5 mL·s-1,刺激时间1.0 s,触发延迟0.5 s。每处理在同一根触角上重复测3次,连续测3根,每只雄虫只取1根触角,测试前后均用溶剂做空白对照。
2 结果与分析 2.1 麻疯树柄细蛾交尾行为及其动态节律 2.1.1 求偶行为雌蛾经过黑暗处理后,接受外界光照的刺激便出现求偶行为。雌蛾求偶一般静止不动,腹部末端产卵器外伸,并向下弯曲,弯曲程度接近直角,此时产卵器顶端淡黄色体晶莹饱满。整个过程中雌蛾保持不动,产卵器偶有伸缩。
雄蛾接受雌蛾散发出的性信息素后,便会出现求偶行为。先是静止蛰伏的雄蛾的触角从贴身平伏开始上举,角度逐渐增大到90°左右,再向左右张开或前伸、摆动,接着雄蛾由静止蛰伏状态经举翅展翅、爬动转为振翅扑动,或者从活动中直接展翅扑动,再发展为持续快速振翅追逐异体,最后,在追逐中腹部频频向左右弯曲摆动,张开抱握器,试图抱握交尾。
当雄虫求偶成功后,雌雄尾部结合在一起通常成180°,雌雄头分别朝向两端,但偶尔其交尾角度也小于180°; 其交尾持续时间从90~200 min不等; 在室内温箱饲养的条件下,其交尾行为发生于经历黑暗处理后的光照初期。
2.1.2 交尾的动态节律从图 1可知,麻疯树柄细蛾交尾百分率在1日龄时达到最高,并且随着日龄的增加,交尾率在降低; 在光照开始的1 h内,交尾率也是最高的。随着光照时间的增加,交尾率也在降低。从图 2可知,麻疯树柄细蛾随着日龄的增加,交尾持续时间也在持续增加。

|
图 1 不同日龄麻疯树柄细蛾的交尾节律 Figure 1 Mating rhythms of S. thraustica adults with different daily age |

|
图 2 同日龄麻疯树柄细蛾的交尾时间(平均值± SD) Figure 2 Time of S. thraustica pairs spending on mating in different daily age (mean ± SD) 不同字母表示差异显著(Duncan's新复极差法,P<0.05),图表下同。 Date with different letters are significantly different(Duncan's multiple range test, P < 0.05). The same below for Fig.and Tab. |
由表 1可知:光照开始的1 h内,在不同光照强度下,成虫均能交配,并随着光照强度的增加交尾率也在增加。表 1再次表明随着成虫日龄的增加,交尾率在降低。从表 2可知:随着光照强度的增加,成虫的交尾持续时间在降低,但是光照强度在500~1 000 lx时,2,3,4,5日龄的成虫交尾持续时间并没有显著差异,特别是4,5日龄的成虫,不同的光照强度对其交尾持续时间没太大影响。表 2还显示:随着日龄的增加,麻疯树柄细蛾交尾持续时间也在持续增加。
|
|
|
|

采用腺体提取技术,以正己烷为溶剂的粗提物引起的雄蛾触角的活性明显高于其他三者,差异显著(P<0.05),以正戊烷为溶剂的粗提物的反应值也明显大于二氯甲烷和乙醚(表 3)。
|
|

从图 3可知: 1日龄雄蛾对1日龄处女雌蛾性信息素反应最强烈,电位值最高,随着雄蛾日龄的增加,对性信息素粗提物的反应逐渐下降。并且腺体提取与空气收集2种方法相比,性腺粗提物产生的EAG反应值较空气收集性信息素高一些。

|
图 3 不同日龄雄蛾对1日龄处女雌蛾性信息素粗提物的EAG反应(平均值± SD) Figure 3 The male EAG response of S. thraustica with different daily age to crude extracts of sex pheromone from the 1-day-old female(mean±SD) |
从图 4可知:雄蛾对性信息素粗提物开始产生反应的最低浓度为10-3 FE·(10 μL)-1,但反应值极小。随着性信息素粗提物浓度的增加,雄蛾的EAG反应值也逐渐增加,在浓度为1 FE·(10 μL)-1的时候,达到最高值。

|
图 4 1日龄雄蛾对不同浓度性信息素粗提物的EAG反应(平均值± SD) Figure 4 EAG response of the 1-day-old male to the different concentration of crude extracts of sex pheromone(mean±SD) |
信息素是昆虫种间或个体间行为通信的一种“化学语言”,鳞翅目雌蛾释放性信息素诱导雄蛾产生一系列行为反应,即“长距离”趋化运动和“近距离”行为反应(杜家纬,1988)。对麻疯树柄细蛾“近距离”行为反应的观察表明,其求偶、交配行为与其他鳞翅目昆虫相似。
关昆虫性信息素产生、释放的时间节律研究表明:大多数昆虫性信息素释放的高峰期由求偶行为来确定,产卵器伸出是释放性信息素的信号(王惠等,1997)。对昆虫交尾节律的观察更能进一步从宏观上了解其释放性信息素的节律。麻疯树柄细蛾在羽化几小时后便能交尾,交尾率也在1日龄时达到最高,随日龄增加,交尾率降低(杜家纬,1988)。
麻疯树柄细蛾雌蛾的日龄影响性信息素的分泌,其主要反映在性信息素腺体的发育成熟程度。雌蛾在羽化当天即能释放性信息素,雄蛾也在1日龄达到性成熟,这可能与其成虫寿命短有一定关系。随日龄增加,其交尾持续时间也增加,这是因为较高日龄成虫的交尾率较低,而增加交尾持续时间在一定程度上增加其竞争能力,这是昆虫的一种适应性生殖策略,这样它们能获得较多的交尾机会。
光照影响麻疯树柄细蛾的交尾行为。在光照开始的第1小时内,交尾率最高,随光照时间增加,交尾率降低。这是因为大部分鳞翅目雌蛾释放性信息素的主要刺激因素是光照周期,雌蛾释放性信息素的行为是受光线强弱刺激控制的。所以当结束暗期开始光照后,成虫只要接受到光信号,就能刺激其发生求偶行为,因此光照初期的交尾率较高; 不同的光照强度,哪怕是很弱的光强信号(1 lx),都能刺激麻疯树柄细蛾发生求偶行为。以上都印证了“成虫多数在清晨交尾”的说法(肖银波等,2008)。不同光强对其交尾节律也有一定影响,随着光照强度增加,交尾率也增加,但当光强达到500与1 000 lx时,1日龄成虫交尾率相差却不大,分别为68.89%和69.40%。这些结果证明麻疯树柄细蛾的求偶交尾有一定趋光性,但当光强达到一定阈值时,其对交尾率影响不大。而在不同光强下,成虫交尾持续时间并没有太大差异,特别是4,5日龄成虫,这说明影响交尾持续时间的重要因素是成虫日龄,而非光强度。
通过EAG试验比较4种不同溶剂的提取效果,弱极性溶剂提取效果较好,以正己烷为最佳。根据2种方法提取性信息素粗提物的EAG数据比较,腺体提取优于空气收集。因为散发在空气中的性信息素含量少,空气收集法无法收集彻底; 而腺体提取是将含有高浓度性信息素的性腺直接置于溶剂中浸泡,避免了性信息素在空气中的损失,提取效果更好。
不同日龄雄虫对性信息素粗提物的EAG反应基本和其交尾节律相一致,1日龄时为最高值,随着雄蛾日龄的增加,无论交尾率还是EAG反应均降低。多数鳞翅目昆虫的求偶交尾行为与性信息素的产生和释放具有一致性(Mazor et al., 2005; Gemeno et al., 2000),麻疯树柄细蛾也基本符合这个规律,这说明雄蛾对性信息素粗提物的反应强弱可以决定交尾成功率的高低(Kanno,1981)。从麻疯树柄细蛾的性信息素剂量反应曲线上可看出本次试验雄蛾对性信息素粗提物的EAG电位值总体偏低。这可能是因为提取过程中性信息素有一定损失,最后得到的粗提物也没有经进一步去杂提纯,所以雄蛾对其EAG反应值自然不高; 也可能是由于不同鳞翅目昆虫雌蛾产生性信息素的量差异悬殊所致,例如粉纹夜蛾(Trichoplusia ni)雌蛾的性信息素含量就比黄地老虎(Agrotis segetum)高出1 000多倍(杜家纬,1988),或许麻疯树柄细蛾雌蛾所含性信息素本来就量微。因此,麻疯树柄细蛾释放性信息素的能力有待进一步研究。
本文结果为进一步研究麻疯树柄细蛾的性信息素奠定了基础,特别是光照对其交尾行为的影响,与大多数蛾类黑暗条件下交尾的情况不同。根据对日龄的研究结果,应用性信息素防治的重点应为1日龄成虫。下一步,应利用气相色谱-触角电位技术及质谱技术确定活性成分的化学结构,以便人工合成并应用于种群的动态监测及诱杀防治等方面。
高文财, 邢飞, 任炳忠. 2005. 昆虫利它素的研究概况[J]. 吉林师范大学学报:自然科学版, (1): 36-38. |
杜家纬. 1988. 昆虫信息素及其应用[M]. 北京: 中国林业出版社: 24-25.
|
陆鹏飞, 乔海莉, 王小平, 等. 2007a. 豆野螟成虫日龄对交尾的影响及雄蛾对性信息素的触角电位反应[J]. 昆虫知识, 44(5): 665-670. |
陆鹏飞, 乔海莉, 王小平, 等. 2007b. 豆野螟成虫行为学特征及性信息素产生与释放节律[J]. 昆虫学报, 50(4): 335-342. |
王惠, 陈竹君, 李科友. 1997. 华北大黑鳃金龟性信息素粗提物的分离及活性组分初探[J]. 西北林学院学报, 12(2): 52-56. |
王秀娟, 熊智, 朱晓琴. 2008. 麻疯树应用研究进展[J]. 西南林学院学报, 28(2): 49-53. |
肖银波, 周建华, 刘艳, 等. 2009. 麻疯树柄细蛾形态和生物学特性观察[J]. 昆虫学报, 52(2): 228-233. |
叶辉. 2000. 光照、温度对纵坑切梢小蠹起飞行为的影响[J]. 昆虫知识, 37(6): 342-344. DOI:10.7679/j.issn.2095-1353.2000.098 |
周建华, 肖银波, 肖育贵, 等. 2008. 四川攀西地区麻疯树人工林有害生物初步研究[J]. 中国森林病虫, (6): 24-26. |
Gemeno C, Haynes K F. 2000. Periodical and age-related variation in chemical communication system of black cutworm moth, Agrotis ipsilon[J]. J Chem Ecol, 26(2): 329-342. DOI:10.1023/A:1005468203045 |
Kanno H. 1981. Mating behaviour of the rice stem borer moth, Chilo suppressalis Walker(Lepidoptera: Pyralidae), 5: Critical illumination intensity for female calling and male sexual response under various temperatures[J]. J Appl Entomol Zool, 16(3): 179-185. DOI:10.1303/aez.16.179 |
Mazor M, Dunkelblum E. 2005. Circadian rhythms of sexual behavior and pheromone titers of two closely related moth species Autographa gamma and Cornutiplusia circumflexa[J]. J Chem Ecol, 31(9): 2153-2168. DOI:10.1007/s10886-005-6082-7 |