 2011, Vol. 47
2011, Vol. 47文章信息
- 冯源恒, 李火根, 王龙强, 边黎明, 姚俊修, 施季森
- Feng Yuanheng, Li Huogen, Wang Longqiang, Bian Liming, Yao Junxiu, Shi Jisen
- 鹅掌楸属树种繁殖性能的遗传分析
- Genetic Dissection for the Reproductive Fitness of Liriodendron Derived from Offsprings of Complete-Diallel Crosses
- 林业科学, 2011, 47(9): 43-49.
- Scientia Silvae Sinicae, 2011, 47(9): 43-49.
-
文章历史
- 收稿日期:2010-11-22
- 修回日期:2010-12-26
-
作者相关文章



2. 广西壮族自治区林业科学研究院 南宁 530001
2. Guangxi Forestry Research Institute Nanning 530001
鹅掌楸属(Liriodendron)现仅存2种,即鹅掌楸(L. chinense)和北美鹅掌楸(L. tulipifera)。1963年,我国著名植物育种家叶培忠教授以鹅掌楸为母本、北美鹅掌楸为父本进行了人工杂交试验,获得了鹅掌楸属的种间人工杂种(南京林产工业学院林学系育种组,1973)。杂种鹅掌楸表现出非常明显的杂种优势,生长旺盛,适应能力强,病虫害少,树形美观,枝叶浓密、生长期长,花色鲜艳(王章荣,2005)。但鹅掌楸属的杂交育种研究工作一直受到亲本繁殖能力低的限制,而繁殖能力低下也是鹅掌楸天然群体濒危的重要原因之一(秦慧贞等,1996)。关于这个问题,许多学者在鹅掌楸胚胎学和交配系统方面开展了大量研究。樊汝汶等(1990)发现鹅掌楸胚发育中存在原胚和胚乳发育不协调的现象,随后尹增芳等(1994; 1997)和秦慧贞等(1996)分别对鹅掌楸雌雄配子败育进行了细致研究,周坚等(1994)对鹅掌楸属植物的花粉品质和花粉管生长进行了研究,这些研究分别证实了雌、雄配子败育是鹅掌楸繁殖能力低下的重要原因。在交配系统研究方面,方炎明等(1994)通过对鹅掌楸天然群体与人工群体的生育力比较认为生育力低的原因可能是花粉限制,黄双全等(1998b)通过对花部数量变异与结实率研究认为可能是资源配量限制了结实率。关于鹅掌楸繁殖能力低下的问题就此形成了“花粉限制(pollen limitation) ”与“资源限制(resource limitation) ” 2种观点。但二者都缺乏从遗传学角度的研究支持。因此开展鹅掌楸属树种交配亲和性及其遗传基础研究对于鹅掌楸的繁殖能力评价、杂交亲本选配及杂种优势利用有重要的理论与现实意义。
双列交配设计可以同时估算一般配合力(GCA)和特殊配合力(SCA)等遗传参数,而且具有统计上的稳定性(Kearsey,1965; Pederson,1972),是非常理想的遗传分析模型,因而在作物和林木遗传育种中得到广泛应用(Verma et al., 2003; Allahgholipour et al., 2006; Kumari et al., 2008; 金国庆等,2008)。本研究以鹅掌楸、北美鹅掌楸各4株成年个体作为交配亲本,在检测各亲本花粉活力基础上,通过全双列交配设计,以各组合子代出苗率作为其繁殖性能指标,进而估算繁殖性能的母本效应、父本效应、父母本交互作用效应、一般配合力与特殊配合力等遗传效应,并分析各类遗传效应的相对重要性大小。同时,利用SSR分子标记技术估算各交配亲本间遗传距离,以探讨亲本遗传距离对交配繁殖性能的影响。
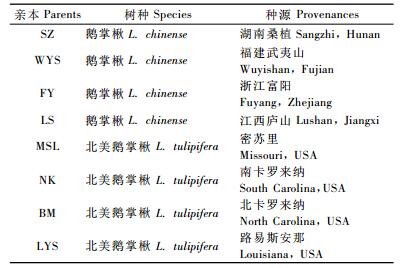
1 材料与方法 1.1 试验材料交配亲本来自江苏句容南京林业大学下蜀实习林场的鹅掌楸属种源试验林。试验林包含12个鹅掌楸种源和5个北美鹅掌楸种源,树龄14年,已进入盛花期和盛果期(李火根等,2005)。2007年5月,在试验林内随机选取4株成年鹅掌楸及4株成年北美鹅掌楸作为交配亲本(表 1)。
|
|
为排除各杂交亲本花粉活力差异影响,控制授粉前对各亲本的花粉活力进行了测定。采集未开放的花苞,每个亲本5 ~ 8朵,要求同一时间段在树冠相同冠层采集; 水培2天后收集花粉,用10%蔗糖+ 50 mg·L-1硼酸+ 1%琼脂培养基培养花粉2.5 h(徐进等,2001); 用显微镜观察花粉萌发状况,每样品重复观察3次,每次观察5个视野,计算萌发率。
1.3 交配设计、控制授粉与杂种培育2007年5月,对8株杂交亲本分别用毛竹搭架,按8 × 8全双列交配设计进行控制授粉。2007年10月采种,共获得63个交配组合的种子(SZ自交组合未收获到种子)。每个组合筛选相对饱满的种子进行出苗率测定。采用完全随机区组设计,4次重复,每重复每组合各100粒翅果,2008年2月播种于南京江浦桥林苗圃,同年5月统计各组合的场圃发芽率。
1.4 交配亲本的SSR分析亲本基因组DNA采用改进的CTAB裂解-硅珠吸附法进行提取(张博等,2004)。紫外分光光度计检测纯度,置于4 ℃冰箱保存备用。
试验所用的引物来自于本实验室从北美鹅掌楸EST序列中开发出的SSR引物(Xu et al., 2010),筛选11对通用性好、多态性高的SSR引物用于本研究。
SSR-PCR反应体系为10 μL,包含20 ~ 24 ng样本DNA,1 × PCR缓冲液(Tris-HCl 10 mmol·L-1 pH 8.0,KCl 50 mmol·L-1),0.26 mmol·L-1 Mg2+,0.2 mmol·L-1 dNTP,正反引物各0.25 μmol·L-1,Taq聚合酶0.25 U(张博等,2004)。扩增反应程序采用Touch-Down PCR: 94 ℃预变性4 min,随后15个Touch-Down循环(94 ℃变性15 s,60 ℃退火15 s,72 ℃延伸30 s,每次循环的退火温度降0.7 ℃),再进入15个主循环(94 ℃变性15 s,49.5 ℃退火15 s,72 ℃延伸30 s),最后20 min延伸反应。SSR-PCR产物采用8%的聚丙烯酰胺变性凝胶电泳,银染检测。对电泳条带判读后,利用PopGen32软件估算亲本间遗传距离(Yeh et al., 2000)。
1.5 数据处理与分析统计各交配组合子代的场圃出苗率作为其繁殖性能指标,计算各亲本繁殖性能的一般配合力(GCA)及各交配组合的特殊配合力(SCA)。鉴于鹅掌楸雌、雄繁殖性能差异较大,所以在估算亲本的GCA时,分别雌、雄繁殖性能进行估算。

|
式中,GCAm为雌性繁殖性能的GCA,gi.为第i个母本的GCAm,xi.为第i个母本的所有组合的均值,GCAf为雄性繁殖性能的GCA,g.j为第j个父本的GCAj,x.j为第j个父本的所有组合的均值,xij为第i个母本和第j个父本交配组合值,u为总平均值(顾万春,2004)。
采用SAS8.1软件,以yijk = μ + Fi + Mj + Dij + eijk为线性模型对父母本效应进行方差分析。式中,yijk为第i个母本和第j个父本交配子代在第k区组的观测值; μ是群体平均值; Fi和Mj分别为母本效应、父本效应; Dij为父母本交互作用效应; eijk为随机误差。
在对父母本效应进行判断后,采用yijk =μ+ gi + gj + sij + rij + eijk线性模型分析GCA,SCA及正反交效应。式中,yijk为第i个母本和第j个父本交配子代在第k区组的观测值; μ是群体平均值; gi和gj为一般配合力效应; sij为特殊配合力效应; rij为正反交效应; eijk为随机误差。
根据方差分析结果进一步估算遗传率。遗传率的计算方法(朱军,1997)为:
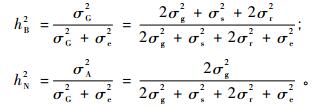
|
式中,hB2,hN2分别为广义遗传率与狭义遗传率,σG2为遗传方差,σA2为加性方差,σg2为一般配合力方差,σs2为特殊配合力方差,σr2为平均亲本效应方差,σe2为环境方差。
2 结果与分析 2.1 鹅掌楸亲本花粉活力检测以亲本的花粉萌发率表示其花粉活力。8株亲本的花粉萌发率(表 2)均较高,在75.1% ~ 96.2%之间。方差分析结果表明,花粉萌发率在8株亲本间差异不显著(F值为0.45,未达0.05差异显著性水平),说明花粉活力对该批亲本繁殖性能的影响可以忽略。
|
|

以各交配组合子代的场圃出苗率作为各组合的繁殖性能。表 3列出了63个交配组合的繁殖性能(场圃发芽率)。63个组合平均繁殖性能为16.26%,其中,鹅掌楸种内交配组合为15.33%,北美鹅掌楸种内交配组合为18.44%,种间杂交组合繁殖性能为15.79%。种间交配组合中,LYS × SZ最高为32.00%,FY × NK最低为0.50%;种内交配组合中,LS × SZ最高为35.50%,FY × WYS和FY × LS最低为0。所有自交组合的繁殖性能均不超过1%。
|
|

进一步对各组合繁殖性能利用SASv8.1软件进行方差分析,结果显示,各交配组合间繁殖性能差异显著,而区组间差异不显著(表 4),表明各杂交组合的繁殖性能差异主要与交配亲本的遗传组成有关。
|
|

由于亲本SZ未获得自交子代,且自交组合育性极低,因此,在估算各亲本繁殖性能配合力时,为了亲本间比较的一致性,仅以56个正反交组合对各杂交亲本的繁殖性能配合力进行估算(表 5)。雌性繁殖性能的一般配合力(GCA) :亲本LYS最高,达11.17; FY最低,仅-14.85。雄性繁殖性能GCA:亲本FY最高,为3.67; LYS最低,为-2.92。平均繁殖GCA最高的是亲本LYS,为4.13;最低的是FY,为-5.59。各亲本间的雌性繁殖GCA差异大于雄性繁殖GCA,同一亲本的雌、雄繁殖性能的GCA差异也较大。
|
|
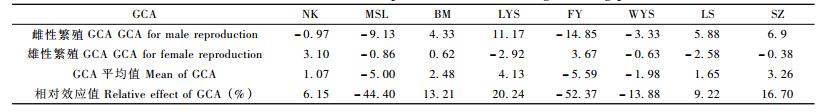
表 6列出了各交配组合繁殖性能的特殊配合力(SCA)。从中可以看出,繁殖性能的SCA在不同交配类型间差异较大,总体上,种内交配的SCA远高于种间杂交。56个正反交组合中,LS × SZ最高,达13.74; LS × NK最低,仅为-10.49。
|
|

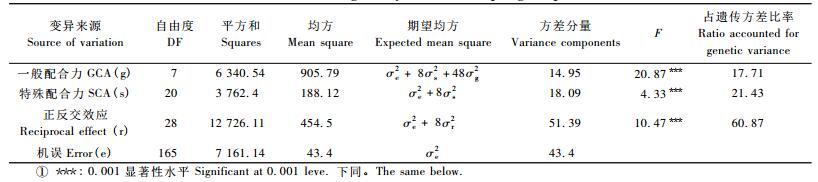
为进一步分析配合力效应对鹅掌楸繁殖性能的影响程度,采用双列杂交方差分析模型(Griffing,1956)对GCA效应、SCA效应和正反交效应的方差分量进行无偏估算。结果表明:对于鹅掌楸繁殖性能,亲本间的GCA效应、SCA效应和正反交效应均达极显著水平; 在总遗传方差中,三者所占比率分别为17.71%,21.43%和60.87% (表 7)。
|
|
为进一步分析影响鹅掌楸繁殖性能的父、母本效应,采用SASv8.1统计软件对各杂交组合繁殖性能进行方差分析(表 8)。从中可以看出,母本效应与交互效应均达到差异显著水平,而父本效应差异不显著。母本效应的方差分量占总遗传方差的71.22%,而父本效应方差分量占总遗传方差的0.01%,这表明鹅掌楸的繁殖性能主要受母本遗传效应影响。
|
|

根据加性方差VA = 2σg2,显性方差VD = σs2,估算出鹅掌楸繁殖性能的加性方差、显性方差分别为29.9,18.9。由亲本效应分析结果可知,鹅掌楸的繁殖性能的父本效应极低,几乎可以忽略不计。因此,可将正反交效应的方差分量作为母本效应,即VF=2σr2。估算出鹅掌楸繁殖性能的广义遗传力为0.776,狭义遗传力为0.154。
2.6 亲本遗传距离对杂交亲和性的影响基于12个SSR位点信息,利用PopGen32软件估算8个杂交亲本间的遗传距离(GD) (表 9)。种间杂交组合中,亲本间遗传距离最大2.853 (SZ与BM),最小0.629(FY与LYS),平均为1.475;种内杂交组合中,亲本间遗传距离最大1.28 (BM与MSL),最小0.405(LYS与MSL),平均为0.816。
|
|
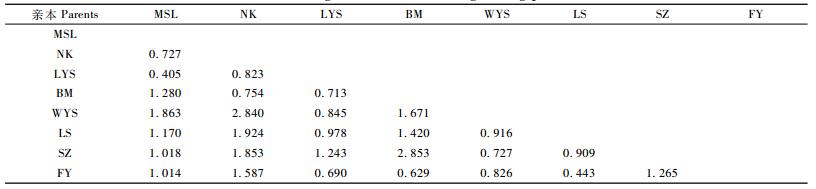
为了解亲本遗传距离与杂交亲和性之间的关系,以各交配组合的繁殖性能特殊配合力作为杂交亲和性的度量指标,将亲本遗传距离与各交配组合的繁殖性能特殊配合力进行相关分析(图 1)。结果显示,总体上,二者相关性不明显(相关系数为-0.09,显著性水平P = 0.52),但也存在一定的趋势:多数种内交配组合双亲的GD值在0.4 ~ 1之间,其SCA随着GD增大而升高(相关系数为0.31,显著性水平P = 0.18);而大多数种间交配组合的GD值在0.6 ~ 2之间,SCA随着GD减小而增大(相关系数为-0.56,显著性水平P = 0.002)。

|
图 1 亲本遗传距离与杂交亲和性的关系 Figure 1 The relationship between genetic distances and cross-compatibility |
评价某种植物的繁殖能力可以从不同的生物学指标入手(Suitor et al., 2009),例如其对繁殖资源的配置投入、产种量或最终成活子代的数量。成年的鹅掌楸配置了大量繁殖资源(李火根等,2007; 冯源恒等,2010),但得到的子代数量却很低(方炎明等,1994),这说明鹅掌楸的繁殖成功率不高。本研究利用鹅掌楸全双列杂交组合子代出苗率作为度量其繁殖性能指标,分析结果显示,鹅掌楸繁殖性能主要决定于母本效应,父本效应非常微弱。由此可见,在对鹅掌楸不同交配组合的繁殖性能进行评价时,应以其雌性繁殖能力作为主要指标。
3.2 鹅掌楸结实率低的原因在以往的研究中,关于影响鹅掌楸自然条件下结实率低的因素一直存在2种观点,即“花粉限制(pollen limitation) ”与“资源限制(resource limitation) ”。方炎明等(1994)通过对鹅掌楸天然群体与人工群体的生育力研究发现饱满翅果数与总翅果数之间不存在负相关,认为鹅掌楸生育力低可能是受花粉限制。采用人工授粉可以明显提高结实率的事实为这一观点提供了有力的支持,但该观点并未进行关于雌配子体败育对结实率影响的讨论。黄双全等(1998b)发现自然传粉的单果结实率也可高达67.4%,对雌性较高的资源投资有利于结实,且鹅掌楸聚合果中存在着种子选择性败育现象,并结合秦慧贞等(1996)关于鹅掌楸胚囊发育率较高的居群其结实率也较高的结论,提出鹅掌楸结实率低的资源限制说(黄双全等,1998a)。本研究获得的鹅掌楸繁殖主要受母本遗传控制这一结论为“胚珠限制”假说提供了有力支持,同时也说明通过选择繁殖性能强的母本可提高鹅掌楸结实率。
3.3 亲本遗传距离对鹅掌楸繁殖性能的影响将亲本遗传距离与表型性状进行相关研究是分子遗传学与常规育种研究的重要结合点(Kiula et al., 2008; Stelkens et al., 2009; Kim et al., 2010)。有研究表明亲本间遗传变异程度可作为亲本选配和子代预测的依据(Jose et al., 2005; 李善文等,2007),但也有研究发现亲本的遗传距离与子代性状虽存在相关性,但其相关程度较低(Kopp et al., 2002; Yu et al., 2005)。本研究将各杂交组合繁殖性能的特殊配合力作为双亲杂交亲和性的度量指标,分析亲本遗传距离与交配组合繁殖性能的特殊配合力二者的相关性。从分析结果来看,虽然总体上,二者相关性不显著,但也可看出一定的趋势。当双亲的遗传距离小于1时,二者杂交亲和性较好,遗传距离大于1时,则杂交亲和性会随之降低。对比各亲本对遗传距离值,不难发现,大多数种内交配组合的亲本间遗传距离小于1,而大多数种间交配组合的亲本间遗传距离大于1。因此,可以初步认为,鹅掌楸种内交配时,杂交亲和性随双亲遗传距离增加而增加; 而种间交配时,杂交亲和性随双亲遗传距离增加而降低。但当双亲遗传距离更大时,如超过2.5的4个组合,则又与此不符。相似的结果在马尾松(Pinus massoniana)和水稻(Oryza sativa)的研究中也有报道,只在一定的遗传距离范围内,或仅对于部分研究材料而言,亲本的遗传距离与子代性状呈显著相关(Zhang et al., 1996; 罗小金等,2006; 张一等,2010)。
但为何在鹅掌楸种内交配时,其杂交亲和性随双亲遗传距离增加而增加,分析其原因,可能是由于鹅掌楸种内杂交可能存在一定程度的近交衰退。亲本遗传距离小、基因相似度高,将导致杂交子代纯合基因位点频率升高、隐性有害基因表达几率增大,从而造成繁殖成功率下降。
3.4 鹅掌楸繁殖性能与育种亲本选择鹅掌楸杂交育种和资源保护一直受到其繁殖能力低的限制,因此,在鹅掌楸育种工作中,除了重视对经济性状的遗传改良外,对繁殖能力的改良也不容忽视。特别是在杂交种子园的建园材料选择时,应重视其繁殖能力。从本研究的结果来看,鹅掌楸繁殖性能所受遗传因素控制的程度依次为:母本效应>特殊配合力效应>一般配合力效应,主要受母本遗传控制。不同的鹅掌楸亲本在雌性繁殖配合力上存在较大变异,说明选择繁殖能力超强的母本是可行的,改良潜力大。此外,在进行鹅掌楸种间杂交育种研究时,选择双亲间遗传距离较近的组合获得较好繁殖效果的可能性更大些。
樊汝汶, 尹增芳, 尤录祥. 1990. 中国鹅掌楸花芽分化的细胞学形态学观察[J]. 南京林业大学学报, 14(2): 26-32. |
方炎明, 尤录祥, 樊汝汶. 1994. 中国鹅掌楸天然群体与人工群体的生育力[J]. 植物资源与环境, 3(3): 9-13. |
冯源恒, 李火根, 杨建, 等. 2010. 两种鹅掌楸繁殖成效的比较[J]. 热带亚热带植物学报, 18(1): 9-14. |
顾万春. 2004. 统计遗传学[M]. 北京: 科学出版社: 292-298.
|
黄双全, 郭友好, 陈家宽. 1998a. 渐危植物鹅掌楸的授粉率及花粉管生长[J]. 植物分类学报, 36(4): 310-316. |
黄双全, 郭友好, 吴艳, 等. 1998b. 鹅掌楸的花部数量变异与结实率[J]. 植物学报, 40(1): 22-27. |
金国庆, 秦国峰, 刘伟宏, 等. 2008. 马尾松生长性状交配效应的遗传分析及杂交组合选择[J]. 林业科学, 44(6): 28-33. DOI:10.11707/j.1001-7488.20080606 |
李火根, 陈龙, 梁呈元, 等. 2005. 鹅掌楸属树种种源试验研究[J]. 林业科技开发, 19(5): 13-16. |
李火根, 曹晓明, 杨建. 2007. 两种鹅掌楸的开花习性与传粉媒介[J]. 浙江林学院学报, 24(4): 401-405. |
李善文, 张志毅, 于志水, 等. 2007. 杨树杂交亲本分子遗传距离与子代生长性状的相关性[J]. 林业科学, 43(1): 35-41. |
罗小金, 贺浩华, 彭小松, 等. 2006. 利用SSR标记分析水稻亲本间遗传距离与杂种优势的关系[J]. 植物遗传资源学报, 7(2): 209-214. |
南京林产工业学院林学系育种组. 1973. 亚美杂种马褂木的育成[J]. 林业科技通讯, (12): 10-11. |
秦慧贞, 李碧媛. 1996. 鹅掌楸雌配子体败育对生殖的影响[J]. 植物资源与环境, 5(3): 1-5. |
王章荣. 2005. 鹅掌楸属树种杂交育种与利用[M]. 北京: 中国林业出版社: 16-34.
|
徐进, 王章荣. 2001. 杂种鹅掌楸及其亲本花部形态和花粉活力的遗传变异[J]. 植物资源与环境学报, 10(2): 31-34. |
尹增芳, 樊汝汶. 1994. 中国鹅掌楸雄配子体发育的超微结构研究[J]. 植物资源与环境, 3(1): 1-8. |
尹增芳, 樊汝汶. 1997. 中国鹅掌楸花粉败育过程的超微结构观察[J]. 植物资源与环境, 6(1): 1-7. |
张博, 张露, 诸葛强, 等. 2004. 一种高效的树木总DNA提取方法[J]. 南京林业大学学报, 28(1): 13-17. |
张一, 储德裕, 金国庆, 等. 2010. 马尾松亲本遗传距离与子代生长性状相关性分析[J]. 林业科学研究, 23(2): 215-220. |
周坚, 樊汝汶. 1994. 鹅掌楸属2种植物花粉品质和花粉管生长的研究[J]. 林业科学, 30(5): 405-411. |
朱军. 1997. 遗传模型分析方法[M]. 北京: 中国农业出版社: 98-111.
|
Allahgholipour M, Ali A J. 2006. Gene action and combining ability for grain yield and its components in rice[J]. Journal of Sustainable Agriculture, 28(3): 39-53. DOI:10.1300/J064v28n03_05 |
Griffing B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems[J]. Aust J Boil Sci, 9(4): 463-493. DOI:10.1071/BI9560463 |
Jose M A, Iban E, Silvia A, et al. 2005. Inheritance mode of fruit traits in melon: heterosis for fruit shape and its correlation with genetic distance[J]. Euphytica, 144(1): 31-38. |
Kearsey M J. 1965. Biometrica1 analysis of a random mating population: a comparison of five experimental designs[J]. Heredity, 20(3): 205-235. |
Kim H S, Kim J G, Baek S B, et al. 2010. Estimation of genetic distance and its predictability of F1 hybrid performance in barley[J]. J Crop Sci Biotech, 13(1): 47-52. DOI:10.1007/s12892-009-0128-2 |
Kiula B A, Lyimo N G, Botha A M. 2008. Association between AFLPbased genetic distance and hybrid performance in tropical maize[J]. Plant Breeding, 127(2): 140-144. DOI:10.1111/j.1439-0523.2007.01434.x |
Kopp R F, Smart L B, Maynard C A, et al. 2002. Predicting withinfamily variability in juvenile height growth of Salix based upon similarity among parental AFLP fingerprints[J]. Theor Appl Genet, 105(10): 106-112. |
Kumari J, Gadag R N, Jha G K. 2008. Combining ability for field emergence, kernel quality traits, and certain yield components in sweet corn (Zea mays L.)[J]. Journal of Crop Improvement, 22(1): 66-81. DOI:10.1080/15427520802043018 |
Pederson D G. 1972. A comparison of four experimental designs for the estimation of heritability[J]. Theor Appl Genet, 42(3): 371-377. |
Stelkens R, Seehausen O. 2009. Genetic distance between species predicts novel trait expression in their hybrids[J]. Evolution, 63(4): 884-897. DOI:10.1111/evo.2009.63.issue-4 |
Suitor S, Potts B M, Brown P H, et al. 2009. The relationship of the female reproductive success of Eucalyptus globulus to the endogenous properties of the flower[J]. Sex Plant Reprod, 22(1): 37-44. DOI:10.1007/s00497-008-0089-5 |
Verma O P, Santoshi U S, Srivastava H K. 2003. Governance of gene action and combining ability for certain grain quality traits in three diverse rice (Oryza sativa L.) growing ecosystems[J]. Journal of Sustainable Agriculture, 22(4): 63-78. DOI:10.1300/J064v22n04_06 |
Xu M, Sun Y, Li H. 2010. EST-SSRs development and paternity analysis for Liriodendron spp[J]. New Forests, 40(3): 361-382. DOI:10.1007/s11056-010-9205-0 |
Yeh F C, Yang R C. 2000. Popgene version 1. 32. [2000-12]. http://www.ualberta.ca/~fyeh/download.htm.
|
Yu C Y, Hu S W, Zhao H X, et al. 2005. Genetic distances revealed by morphological characters, isozymes, proteins and RAPD markers and their relationships with hybrid performance in oilseed rape (Brassica napus L).[J]. Theor Appl Genet, 110(3): 511-518. DOI:10.1007/s00122-004-1858-7 |
Zhang Q F, Zhou Z Q, Yang G P, et al. 1996. Molecular marker heterozygosity and hybrid performance in indica and japonica rice[J]. Theor Appl Genet, 93(8): 1218-1224. DOI:10.1007/BF00223453 |