 2011, Vol. 47
2011, Vol. 47文章信息
- 铁军, 张晶, 彭林鹏, 赵本元, 张志翔
- Tie Jun, Zhang Jing, Peng Linpeng, Zhao Benyuan, Zhang Zhixiang
- 夏秋季节神农架川金丝猴取食主要影响因素分析
- Analysis of Main Factors Influencing Summer and Autumn Feeding of Rhinopithecus roxellana in Shennongjia Nature Reserve
- 林业科学, 2011, 47(7): 108-115.
- Scientia Silvae Sinicae, 2011, 47(7): 108-115.
-
文章历史
- 收稿日期:2009-11-30
- 修回日期:2011-03-20
-
作者相关文章



2. 北京林业大学生物科学与技术学院 北京 100083;
3. 湖北神农架自然保护区管理局 神农架 442421;
4. 国家林业局自然保护区研究与评价中心 北京 100083
2. College of Biological Sciences and Technology, Beijing Forestry University Beijing 100083;
3. Management Bureau of Hubei Shennongjia National Nature Reserve Shennongjia 442421;
4. Nature Reserve Research and Assessment Center of State Forestry Administration Beijing 100083
川金丝猴(Rhinopithecus roxellana)是中国特有的珍稀物种,国家Ⅰ级保护动物,主要分布于陕西、四川、甘肃及湖北等地(汪松等,1998; 全国强等,2002)。湖北神农架是川金丝猴分布的最东端(张荣祖等,1997),在保护区内主要分布于金猴岭和千家坪地区(朱兆泉,2003),现有8群约1 200只。
食源植物中蛋白质、脂肪等营养物质和次生化合物含量的时空变化对野生动物的食性选择有一定的影响(Nolte et al., 1992; Nagy et al., 1964; Radwan et al., 1974; Robbins et al., 1987)。对于树栖的川金丝猴而言,除了栖息地植物构成和地形因素外,食源植物的丰富度、含水量、可消化能量、营养成分含量、次生代谢产物种类及含量等因素也会对川金丝猴的食物选择产生一定的影响。本文针对夏、秋季节神农架自然保护区内川金丝猴的食物组成和主要食源植物丰富度、营养成分进行分析,旨在揭示川金丝猴食性选择规律,了解神农架川金丝猴的夏秋季节取食对策,以期为川金丝猴的保护和管理提供基本资料和依据。
1 研究地概况与研究方法 1.1 研究地基本概况神农架自然保护区地处湖北省西北部(109°56'—110°58' E,30°15'—31°57' N),以秦巴山脉东端的神农架山系构成保护区主体,山体大致呈东西走向,最高峰神农顶海拔3 105.4 m,为中国地势第二阶梯向第三阶梯的过渡区域。神农架地跨中、北亚热带,深受东南、西南季风影响,气候温暖湿润,具有明显的季节性,年均温度约12 ℃,年均降水量在1 800 mm以上; 春季较短暂,从4月至5月底,夏季一般从6月到8月中旬,7月份平均温度约23 ℃,秋季从8月中旬到10月底,凉爽少雨,而冬季较漫长,从11月延续到次年的3月底,12月到2月底有雪覆盖,其中1月份的温度最低,平均温度约-2.5 ℃ (朱兆泉等,1999)。独特的地理位置及受第四纪冰川的影响较小,造就了这里植物物种的丰富性与古老性,是我国生物多样性保护关键区域之一(葛继稳等,1997)。
1.2 研究方法 1.2.1 川金丝猴食源植物种类调查2006—2008年,对神农架国家自然保护区千家坪地区(110°03'—110°34' E,31°22'—31°37' N,距木鱼镇约13 km)川金丝猴活动范围内的主要植被类型进行了8次实地调查。在栖息地海拔1 800~2 600 m之间,设置8个海拔梯度(约每隔100 m设1条样带),20 m × 20 m的样方255个,其中夏季设置125个,秋季设置130个。具体调查内容见铁军等(2009)。
1.2.2 川金丝猴食源植物资源谱的测定记录样方内食源植物的株数,测量株高(目测求平均),生物量采用获取法,按2 cm为径级选取径级标准木,每个径级选取5株。选定标准木后,以2 m区分段,用分层切割法取食源植物的嫩枝(条)、叶片和果实等可食部分,采集不同器官鲜质量样品500 g在80 ℃恒温下烘干至恒重,测量不同器官的干物质质量。统计所测样方中的数据,求出各种食源植物的平均高度、平均生物量和平均频度,并计算出样方内各种食源植物的相对高度(relative height,RH)、相对生物量(relative biomass,RB)和相对频度(relative frequency,RF)。具体计算公式及内容见铁军等(2010)。
1.2.3 植物种类的识别及镜检植物样本和粪便样品处理及样片制备参照吴建平等(2005)和崔多英等(2007)。
在100倍的显微镜下,观察各种植物样片中植物角质表皮细胞的形状、大小、排列方式以及细胞壁的厚度、气孔的形状、大小及密度; 表皮毛基细胞的大小、形状及密度等可鉴别的形态特征,并用显微照相系统对各种植物进行数码拍照,作为食性分析时显微鉴定的对照样本。在观察粪便样片时,每个样片随机选择20个视野,寻找植物的鉴定特征(参照植物显微照片),镜检过程中,统计每个粪便样片中显微镜视野范围内各种植物表皮碎片的总数及单种植物表皮碎片数。计算出每种植物在动物粪便中的相对丰富度(relative density,RD),即动物粪便中的食物组成(曹伊凡等,2008),公式为:
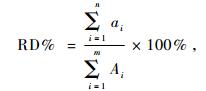
|
式中: ai表示植物在第i个粪样中植物碎片数; Ai表示植物在第i个粪样中总植物碎片数,i = 1,2,3,…,n。
1.2.4 食源植物营养成分的测定夏秋季节采集川金丝猴主要食源植物可食部分,用天平称取植物样品的鲜质量,每种植物采集量约1 000 g,自然风干后封存于布袋内。在实验室内,于60 ℃烘干48 h后粉碎并称量,然后过筛(40目),袋装密封待测。
采用概略养分分析法(feed proximate analysis)测定各食源植物的水分(water,W)、粗蛋白(crude protein,CP)、粗脂肪(ether extract,EE)、粗灰分(crude ash,CA)及粗纤维(crude fiber,CF)含量,并间接计算出无氮浸出物(nitrogen-free extract,NFE)的含量,公式为:
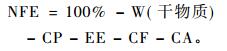
|
食源植物粗蛋白、粗脂肪和粗纤维含量测定在中国农业科学院饲料所中心分析实验室完成; 粗灰分、水分等含量测定在北京林业大学生物学院中心分析实验室完成。
1.2.5 数据统计与处理利用Excel 2003和SPSS 16.0软件对神农架川金丝猴食物组成(RD)与影响其取食的7种因素:食源植物相对丰富度(relative abundance,RA)、食源植物中的水分、粗蛋白、粗脂肪、粗纤维、粗灰分、无氮浸出物含量数据进行相关性分析、T检验和F检验,并对上述7种因素进行主成分分析,确定夏秋季节川金丝猴采食的主要影响因子。
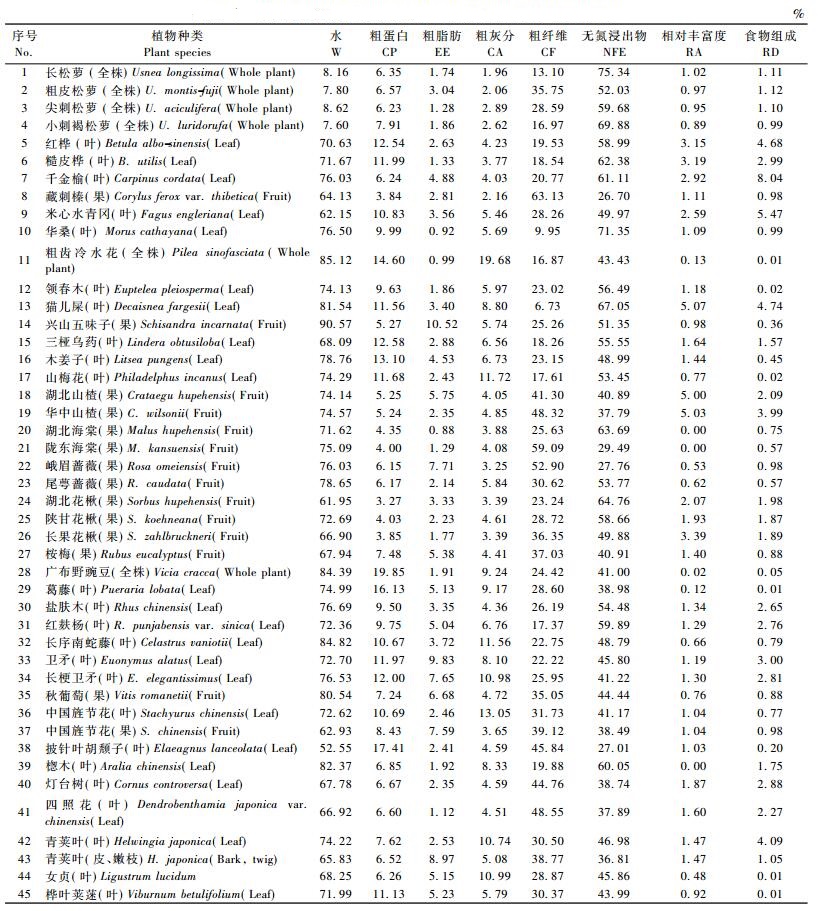
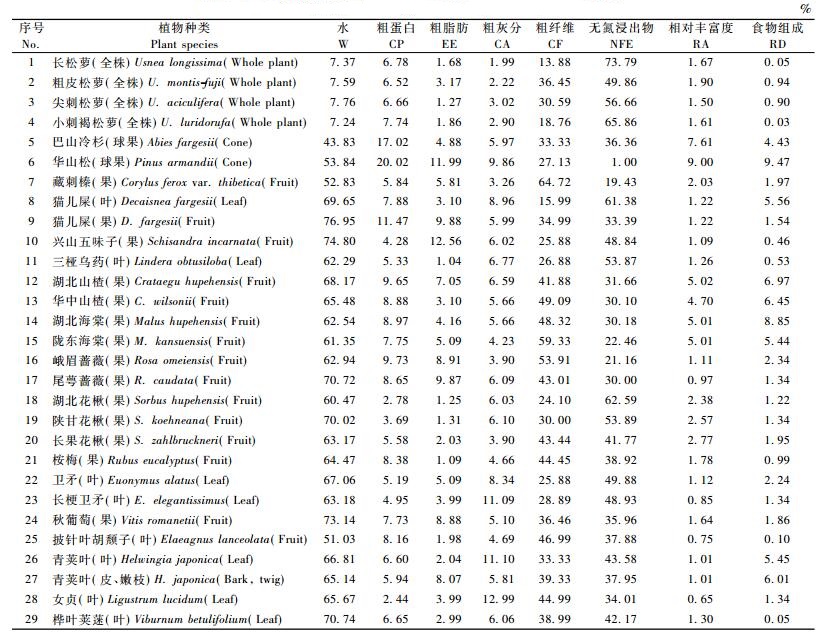
2 结果与分析 2.1 夏秋季节川金丝猴食物组成与食源植物丰富度的关系研究发现:神农架川金丝猴夏季和秋季主要采食的植物有45种,可食部位有48个,其中,夏季主要采食的植物有43种,45个部位; 秋季27种,29个部位(表 1,2)。
|
|
|
|
对神农架川金丝猴夏秋季节主要食物组成与食源植物相对丰富度之间的相关性进行分析,结果表明:川金丝猴食物组成与食源植物相对丰富度,在夏季(n = 45,r = 0.656,P = 0.000)和秋季(n = 29,r = 0.703,P = 0.000)均呈正相关; 食物组成与食源植物相对丰富度在夏季(F = 0.264,df = 87,P = 0.609)和秋季(F = 0.372,df = 57,P = 0.544)差异均不显著(P>0.05)。
2.2 夏秋季节川金丝猴取食与食源植物营养成分的关系 2.2.1 川金丝猴取食与食源植物含水量的关系食源植物含水是川金丝猴获取水分的主要来源。不同季节川金丝猴采食不同种类的植物,获取水分的多少也不尽相同,食源植物含水量可能成为食物质量的一项重要指标。从夏秋季节川金丝猴食源植物不同部位的平均水分含量来看(表 1,2),除4种松萝的含水量夏秋季节呈无明显变化外,夏季含水量最高的是兴山五味子(果),达90.57%,粗齿冷水花(全株)、长序南蛇藤(叶)、广布野豌豆(全株)、楤木(叶)和猫儿屎(叶)次之,分别为85.12%,84.82%,84.39%,82.37%和81.54%,披针叶胡颓子(叶)最低,为52.55%;秋季含水最高的是猫儿屎(果),为76.95%,其次为兴山五味子(果)、秋葡萄(果)、桦叶荚蒾(叶)、尾萼蔷薇(果)和陕甘花楸(果),分别为74.80%,73.14%,70.74%,70.72%和70.02%,巴山冷杉(球果)最低,为43.83%。
对神农架夏秋季节川金丝猴食源植物含水量与其食物组成的相关分析可知:夏季(n = 45,r = 0.078,P = 0.609)和秋季(n = 29,r = 0.258,P = 0.176)食物组成与食源植物含水量呈正相关; 食物组成与食源植物含水量在夏季(F = 481.322,df = 89,P = 0.000)和秋季(F = 183.574,df = 57,P = 0.000)差异均极显著(P<0.01)。
2.2.2 川金丝猴取食与食源植物粗蛋白的关系蛋白质是野生动物生命最基本的营养物质,在动物的生长、发育、繁殖中起着重要作用。从表 1,2可知:夏季,食源植物中粗蛋白含量最高的是广布野豌豆(全株),达19.85%,披针叶胡颓子(叶)、葛藤(叶)、粗齿冷水花(全株)、木姜子(叶)和三桠乌药(叶)次之,分别为17.41%,16.13%,14.60%,13.10%和12.58%,湖北花楸(果)的含量最低,为3.27%;秋季,粗蛋白含量最高的是华山松(球果),为20.02%,其次为巴山冷杉(球果)、猫儿屎(果)、峨眉蔷薇(果)和湖北山楂(果)、湖北海棠(果),分别为17.02%,11.47%,9.73%,9.65%和8.97%,女贞(叶)的含量最低,为2.44%。
从食物组成与食源植物粗蛋白含量的相关性分析可知:川金丝猴食物组成与食源植物粗蛋白含量在夏季(n = 45,r =-0.113,P = 0.459)呈负相关,秋季(n = 29,r = 0.537,P = 0.003)呈正相关; 但夏季(F = 127.858,df = 89,P = 0.000)和秋季(F = 31.908,df = 57,P = 0.000)川金丝猴食物组成与食源植物粗蛋白含量差异均极显著(P<0.01)。
2.2.3 川金丝猴取食与食源植物粗纤维含量的关系植物纤维素含量的高低是影响野生动物食性选择的重要因素,粗纤维含量直接决定可消化能量的高低。从表 1,2可知:在夏季,藏刺榛(果)粗维素含量是食源植物中最高的,达63.13%,陇东海棠(果)、峨眉蔷薇(果)、四照花(叶)、华中山楂(果)和披针叶胡颓子(叶)次之,分别为59.09%,52.90%,48.55%,48.32%和45.84%,猫儿屎(叶)的含量最低,为6.73%;在秋季,食源植物粗维素含量最高的仍为藏刺榛(果),为64.72%,其次为陇东海棠(果)、峨眉蔷薇(果)、华中山楂(果)、湖北海棠(果)和披针叶胡颓子(叶),其含量分别为59.33%,53.91%,49.09%,48.32%和46.99%,长松萝(全株)的含量最低,为13.88%。
对食物组成与粗纤维含量的相关性分析可知:川金丝猴主要食物组成与食源植物粗纤维含量在夏季(n = 45,r =-0.144,P = 0.347)呈负相关,秋季(n = 29,r = 0.194,P = 0.312)则呈正相关; 但夏季(F = 219.301,df = 89,P = 0.000)和秋季(F = 207.154,df = 57,P = 0.000)食物组成与食源植物粗纤维含量差异均极显著(P<0.01)。
2.2.4 川金丝猴取食与食源植物粗灰分含量的关系从表 1,2可知:在夏季,粗齿冷水花(全株)粗灰分含量在食源植物中最高,为19.68%,中国旌节花(叶)、山梅花(叶)、长序南蛇藤(叶)、女贞(叶)和长梗卫矛(叶)次之,分别为13.05%,11.72%,11.56%,10.99%和10.98%,长松萝(全株)的含量最低,为1.96%;在秋季,女贞(叶)的含量最高,为12.99%,青荚叶(叶)、长梗卫矛(叶)、华山松(球果)、猫儿屎(叶)和卫矛(叶)次之,分别为11.10%,11.09%,9.86%,8.96%和8.34%,长松萝(全株)的含量最低,为1.99%。
对食物组成与食源植物粗灰分含量的相关性分析可知:川金丝猴主要食物组成与粗灰分含量在夏季(n = 45,r =-0.156,P = 0.305)呈负相关,秋季(n = 29,r = 0.308,P = 0.104)呈正相关; 但夏季(F = 58.466,df = 89,P = 0.000)和秋季(F = 20.390,df = 57,P = 0.000)的食物组成与食源植物粗灰分含量差异均极显著(P<0.01)。
2.2.5 川金丝猴取食与食源植物粗脂肪含量的关系从表 1,2可知:在夏季,兴山五味子(果)的粗脂肪含量最高,为10.52%,卫矛(叶)、青荚叶(皮、嫩枝)、峨眉蔷薇(果)、长梗卫矛(叶)和中国旌节花(果)次之,分别为9.83%,8.97%,7.71%,7.65%和7.59%,湖北海棠(果)的含量最低,为0.88%;在秋季,仍为兴山五味子(果)的粗脂肪含量最高,为12.56%,华山松(球果)、猫儿屎(果)、尾萼蔷薇(果)、峨眉蔷薇(果)和秋葡萄(果)次之,分别为11.99%,9.88%,9.87%,8.91%和8.88%,三桠乌药(叶)的粗脂肪含量最低,为1.04%。
对食物组成与粗脂肪含量的相关性分析可知:川金丝猴主要食物组成与食源植物粗脂肪在夏季(n = 45,r = 0.020,P = 0.896)和秋季(n = 29,r = 0.291,P = 0.125)均呈正相关; 川金丝猴主要食物组成与食源植物粗脂肪于夏季(F = 19.593,df = 89,P = 0.000)差异极显著(P<0.01),而秋季(F = 5.729,df = 57,P = 0.020)差异显著(p<0.05)。
2.2.6 川金丝猴取食与食源植物无氮浸出物含量的关系无氮浸出物主要包括淀粉、糖类和有机酸,是动物营养中的重要能量来源。从表 1,2可知:在夏季,长松萝(全株)的无氮浸出物含量最高,为75.34%,华桑(叶)、小刺褐松萝(全株)、猫儿屎(叶)、湖北花楸(果)和湖北海棠(果)为次之,分别为71.35%,69.88%,67.05%,64.76%和63.69%,藏刺榛(果)的含量最低,为26.70%;在秋季,仍为长松萝(全株)的无氮浸出物含量最高,为73.79%,小刺褐松萝(全株)、湖北花楸(果)、猫儿屎(叶)、尖刺松萝(全株)和陕甘花楸(果)次之,分别为65.86%,62.59%,61.38%,56.66%和53.89%,华山松(球果)的含量最低,为1.00%。
对食物组成与无氮浸出物含量的相关性分析可知:夏季(n = 45,r = 0.238,P = 0.115)川金丝猴食物组成与食源植物无氮浸出物呈正相关,秋季(n = 29,r =-0.546,P = 0.002)则呈负相关; 川金丝猴食物组成与食源植物无氮浸出物含量于夏季(F = 687.704,df = 89,P = 0.000)和秋季(F = 168.791,df = 57,P = 0.000)差异均极显著(P<0.01)。
通过对夏秋季节可供采食的川金丝猴食源植物(共26种采食部位)营养成分进行配对T检验,发现夏秋季节川金丝猴食源植物的含水量(t = 6.321,df = 25,P = 0.000)、粗灰分含量(t =-4.487,df = 25,P = 0.000)、粗纤维含量(t =-4.079,df = 25,P = 0.000)和无氮浸出物含量(t = 3.045,df = 25,P = 0.005)差异均极显著(P<0.01);而食源植物的粗蛋白含量(t = 0.894,df = 25,P = 0.380)、粗脂肪含量(t =-0.189,df = 25,P = 0.851),食源植物相对丰富度(t =-1.283,df = 25,P = 0.211)和食物组成(t =-2.221,df = 25,P = 0.036)差异均不显著(P>0.05)。
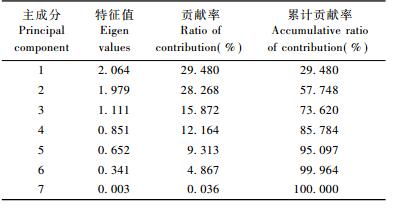
2.3 夏秋季节川金丝猴取食相关因子的主成分分析 2.3.1 夏季川金丝猴取食相关因子的主成分分析对影响川金丝猴夏季取食的7项因子(食源植物的含水量、粗蛋白、粗纤维、粗脂肪、粗灰分、无氮浸出物、相对丰富度)进行主成分分析,可以看出:特征值大于1的主成分有3个,累计贡献率达73.62% (表 3),说明前3个主成分包含了川金丝猴食源植物含水分、粗蛋白、粗纤维和粗脂肪等7个变量的大部分信息,其余4个成分对方差影响很小。因此,取前3个主成分进行进一步分析计算出其相应的特征向量,找出影响川金丝猴夏季取食的主要因子(表 4)。
|
|
|
|

从表 3,4可以看出:第一主成分的贡献率较高(29.48%),反映了夏季川金丝猴取食的主要方面,特征向量中食源植物的粗灰分、粗蛋白、水分和相对丰富度的相关系数绝对值大,分别为0.870,0.776,0.475和0.466,表明食源植物的粗灰分、粗蛋白、水分和相对丰富度等因子对川金丝猴取食有较大影响; 第二主成分的贡献率为28.27%,特征向量中食源植物的无氮浸出物、粗纤维、相对丰富度和粗脂肪的相关系数绝对值大,分别为0.936,0.886,0.362和0.307,反映了食源植物的无氮浸出物、粗纤维、相对丰富度和粗脂肪对川金丝猴取食的影响; 第三主成分的贡献率为15.87%,特征向量中食源植物的水分、相对丰富度、粗脂肪、无氮浸出物的相关系数绝对值较大,分别为0.688,0.638,0.553和0.211,反映了食源植物的水分、相对丰富度、粗脂肪、无氮浸出物对川金丝猴取食的影响。
综合3个主成分的荷载系数值来看,荷载系数值从大到小依次为无氮浸出物(0.936)、粗纤维(0.886)和粗灰分(0.870),表明食源植物的无氮浸出物、粗纤维和粗灰分为夏季神农架川金丝猴取食的主要影响因子。
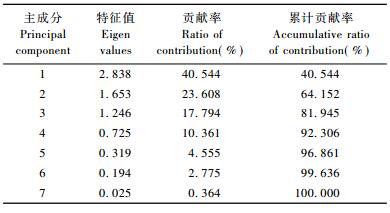
2.3.2 秋季川金丝猴取食相关因子的主成分分析对川金丝猴秋季取食各项相关因子进行主成分分析可知:特征值大于1的主成分有3个,累计贡献率达81.95% (表 5),说明前3个主成分包含了秋季川金丝猴食源植物的含水分量、粗蛋白、粗纤维和粗脂肪等7个变量的大部分信息。因此,取前3个主成分进行进一步分析计算出其相应的特征向量,找出影响川金丝猴秋季取食的主要因子(表 6)。
|
|
|
|
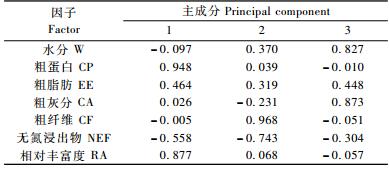
从表 5,6可以看出:第一主成分的贡献率较高(40.54%),反映了秋季川金丝猴取食的主要方面,特征向量中食源植物的粗蛋白、相对丰富度、无氮浸出物和粗脂肪的相关系数绝对值大,分别为0.948,0.877,0.558和0.464,表明食源植物的粗蛋白、相对丰富度、无氮浸出物和粗脂肪等因子对川金丝猴取食有较大影响; 第二主成分的贡献率为23.61%,特征向量中食源植物的粗纤维、无氮浸出物、水分和粗脂肪的相关系数绝对值大,分别为0.968,0.743,0.370和0.319,反映了食源植物的粗纤维、无氮浸出物、水分和粗脂肪对川金丝猴取食的影响; 第三主成分的贡献率为17.79%,特征向量中食源植物的粗灰分、水分、粗脂肪、无氮浸出物的相关系数绝对值较大,分别为0.873,0.827,0.448和0.304,反映了食源植物的粗灰分、水分、粗脂肪、无氮浸出物对川金丝猴取食的影响。
综合来看,荷载系数值从大到小依次为粗纤维(0.968)、粗蛋白(0.948)和食源植物相对丰富度(0.877)。因此,粗纤维、粗蛋白和食源植物相对丰富度为秋季神农架川金丝猴取食的主要影响因子。
3 结论与讨论 3.1 夏秋季节川金丝猴取食与营养成分的关系一般来讲,随着可供采食植物种类的增多,野生动物对食物的选择性也增强(孙儒泳等,1993)。铁军等(2009)研究表明:川金丝猴的食物选择与栖息地植物种群特征密切相关。通过野外啃食调查和粪便显微组织学分析,川金丝猴的食物组成与各种食源植物在群落中的相对生物量、密度和高度呈显著的相关性(铁军等,2010)。本研究发现:神农架川金丝猴食物组成与食源植物相对丰富度夏等(r = 0.656,P = 0.000)和秋季(r = 0.703,P = 0.000)均呈正相关关系。
从川金丝猴主要食源植物营养成分分析可知:神农架内川金丝猴食源植物含水量在夏季为90.57%~52.55%,秋季76.95%~43.83%;粗蛋白含量夏季为19.85%~3.27%,秋季为20.02%~2.44%;食源植物粗维素含量夏季63.13%~6.73%,秋季64.72%~13.88%;食源植物粗灰分含量夏季19.68%~1.96%,秋季12.99%~1.99%;食源植物粗脂肪含量夏季10.52%~0.88%,秋季12.56%~1.04%;食源植物无氮浸出物含量夏季75.34%~26.70%,秋季73.79%~1.00%。对食源植物营养成分与川金丝猴取食食物组成进行相关性分析可知:川金丝猴食物组成与食源植物含水量在夏季和秋季呈正相关; 与食源植物粗蛋白含量于夏季呈负相关,秋季呈正相关; 与食源植物粗纤维量在夏季呈负相关,秋季则呈正相关; 与粗灰分含量在夏季呈负相关,秋季呈正相关; 与食源植物粗脂肪含量在夏秋季节均呈正相关; 与食源植物无氮浸出物含量在夏季呈正相关,秋季则呈负相关。这说明:从春季至冬季,随着气候的不断改变神农架川金丝猴栖息地内的可供采食的源植物种类、取食部位及所含营养成分也不断发生改变,致使川金丝猴的食物组成也随之改变(铁军等,2009; 2010)。
通过对夏秋季节可供采食的川金丝猴食源植物(共26种采食部位)营养成分进行配对T检验,发现夏秋季节川金丝猴食源植物营养质量随着季节的变化,导致川金丝猴的食物组成也发生相应的改变。
3.2 夏秋季节影响川金丝猴取食的主要因子植物的营养质量与植物的物候期有关,其当年植物枝、叶在冬季含有最少的粗蛋白和最多的粗纤维,冬季植物的营养质量下降到最低点(Nelson,1982)。通常情况,果实的含糖量较高,而蛋白质的含量较叶片、种子、茎都要低; 叶片的蛋白质含量较高,纤维素比果实高而比茎低; 种子的蛋白含量质介于果和叶之间(Nakagawa,1989)。植物叶片和果实中的脂类是由半乳糖脂和混杂脂类(色素、角质、蜡、香精油等)组成。半乳糖脂所占比例随着植物的生长发育逐渐减少,而混杂脂类的比例相应增高(郝正里等,2002)。有报道认为:自由采食的野生动物根据自身的营养需要选择和摄取食物,因而食源植物营养成分的种类及含量影响着动物的取食选择(李绍文,1991; Nolte et al., 1992; 王钦等,2002)。灵长类对食物具有高度选择性,通常根据营养选择特定植物的特定部位。只有这样,灵长类才能适应植物界的复杂性(Waterman et al., 1981)。
不同灵长类动物不同个体对食物中所含有的矿质物质及其他营养物质具有不同的选择性(蔡湘文,2004)。从夏秋季节神农架川金丝猴取食各相关因子的主成分分析可以看出:食源植物的无氮浸出物、粗纤维和粗灰分为夏季川金丝猴食物选择的主要影响因子,而食源植物的粗纤维、粗蛋白和相对丰富度为秋季川金丝猴取食的主要影响因子。从本次调查结果来看,川金丝猴的食物选择与黔金丝猴(Rhinopithecus brelichi)和白头叶猴(Trachypithecus leucocephalus)的食物选择较一致,它们均喜欢选择高水分、高蛋白质和低纤维含量的植物嫩叶、嫩枝、果实和种子为食(黄乘明等,1998),而与取食高纤维植物叶片为食的黑叶猴(Presbytis francoisi)不同(黄乘明等,1998; 蔡湘文,2004)。猴类一般喜食含高蛋白质和低纤维和低次代谢生物质的植物叶(McKey et al., 1981; Milton,1979; Oates et al., 1980),对于食源植物所含的次生代谢物质如何影响川金丝猴的取食选择还有待研究。
蔡湘文. 2004. 黑叶猴的觅食生物学和营养分析. 桂林: 广西师范大学生命科学学院硕士学位论文, 39-44. http: //cdmd. cnki. com. cn/article/cdmd-10602-2004130266. htm
|
曹伊凡, 苏建平, 连新明, 等. 2008. 可可西里自然保护区藏羚羊的食性分析[J]. 兽类学报, 28(1): 14-19. |
崔多英, 刘振生, 王小明, 等. 2007. 贺兰山马鹿冬季食性分析[J]. 动物学研究, 28(4): 383-388. |
葛继稳, 吴金清, 朱兆泉, 等. 1997. 神农架生物圈保护区植物多样性及其保护现状的研究[J]. 武汉植物研究, 15(4): 341-352. |
郝正里, 张学炎, 郭艳丽, 等. 2002. 兴隆山自然保护区马麝主要可食野生植物的营养成分[J]. 中国草食动物, 22(4): 41-44. |
黄乘明, 韦毅. 1998.论白头叶猴的食物与营养∥胡锦矗.脊椎动物资源及保护.成都:四川科学技术出版社, 101-105.
|
李绍文. 1991. 生态生物化学(九) :哺乳动物对食物的选择[J]. 生态学杂志, 10(2): 65-71. |
全国强, 谢家骅. 2002. 金丝猴研究[M]. 上海: 上海科技教育出版社: 79-102.
|
孙儒泳, 李博, 诸葛阳. 1993. 普通生态学[M]. 北京: 高等教育出版社: 135-138.
|
铁军, 张晶, 彭林鹏, 等. 2009. 神农架川金丝猴栖息地优势树种生态位及食源植物[J]. 植物生态学报, 33(3): 482-491. |
铁军, 张晶, 彭林鹏, 等. 2010. 神农架川金丝猴冬春季节食性分析[J]. 生态学杂志, 29(1): 62-68. |
汪松, 杨朝飞, 郑光美. 1998. 中国濒危动物红皮书(兽类)[M]. 北京: 科学出版社: 65-68.
|
王钦, 李峻成, 安玉锋, 等. 2002. 放牧生态系统草畜间供与求的动态特征[J]. 草地学报, 10(3): 173-178. DOI:10.11733/j.issn.1007-0435.2002.03.004 |
吴建平, 单继红, 王志平. 2005. 小兴安岭通河林区斑羚冬季食性分析[J]. 动物学杂志, 40(4): 40-44. |
张荣祖, 金善科, 全国强, 等. 1997. 中国哺乳动物分布[M]. 北京: 中国林业出版社: 61.
|
朱兆泉, 宋朝枢. 1999. 神农架自然保护区科学考察集[M]. 北京: 中国林业出版社: 107-121.
|
朱兆泉. 2003. 神农架金丝猴生态学研究[J]. 湖北林业科技, (增): 46-52. |
McKey D B, Gartlan J S, Waterman F L S, et al. 1981. Food selection by black colobus monkeys (Colobus satanas) in relation to food chemistry[J]. Biological Journal of the Linnean Society, 16(2): 115-146. DOI:10.1111/bij.1981.16.issue-2 |
Milton K. 1979. Factors influencing leaf choice by howler monkeys: a test of some hypotheses of food selection by generalist herbivores[J]. American Naturalist, 114(3): 362-378. DOI:10.1086/283485 |
Nagy J G, Steinhoff H W, Ward G M. 1964. Effects of essential oils of sagebrush on deer rumen microbial function[J]. Journal of Wildlife Management, 28(4): 785-790. DOI:10.2307/3798795 |
Nakagawa N. 1989. Bioenergetics of Japanese monkeys(Macaca fuscata) on Kinkazan Island during winter[J]. Primates, 30(4): 441-460. DOI:10.1007/BF02380873 |
Nelson J R. 1982. Nutritional requirement and food habits//Thomas J W, Toweill D E. Elk of North America: Eology and Management. Harrisburg, PA: Stackpole Books, 698. https://core.ac.uk/display/19672358
|
Nolte D I, Provenza F D. 1992. Food preferences in lambs after exposure to flavors in solid foods[J]. Applied Animal Behaviour Science, 32(4): 337-347. DOI:10.1016/S0168-1591(05)80026-7 |
Oates J F, Waterman P G, Choo G M. 1980. Food selection by the south Indian leaf monkey, Presbytis johnii, in relation to leaf chemistry[J]. Oecologia, 45(1): 45-56. DOI:10.1007/BF00346706 |
Radwan M A, Crouch G L. 1974. Plant characteristics related to feeding preference by black-tailed deer[J]. Journal of Wildlife Management, 38(1): 32-41. DOI:10.2307/3800197 |
Robbins C T, Hanley T A, Hagerman A E, et al. 1987. Role of tannins in defending plants against ruminants: reduction in protein availability[J]. Ecology, 68(1): 98-107. DOI:10.2307/1938809 |
Waterman P G, Choo G M. 1981. The effects of digestibility-reducing compounds in leaves on food selection by some Colobinae[J]. Malaysian Applied Biology, 10(2): 147-162. |