 2011, Vol. 47
2011, Vol. 47文章信息
- 丁访军, 王兵, 赵广东
- Ding Fangjun, Wang Bing, Zhao Guangdong
- 毛竹树干液流变化及其与气象因子的关系
- Sap Flow Changes of Phyllostachys edulis and Their Relationships with Meteorological Factors
- 林业科学, 2011, 47(7): 73-81.
- Scientia Silvae Sinicae, 2011, 47(7): 73-81.
-
文章历史
- 收稿日期:2010-11-29
- 修回日期:2011-01-19
-
作者相关文章



2. 贵州省林业科学研究院 贵阳 550005
2. Guizhou Academy of Forestry Guiyang 550005
林分蒸腾量在森林水分循环中占很大的比例(周晓峰,1999)。蒸腾耗水量是树木生理水分的重要参数,如何完整、准确地测定和掌握整株树木的耗水量和耗水规律,并根据树种的耗水特性进行树种选择,合理地构建森林植被,已成为当前生态环境建设的关键。由于树木蒸腾作用受多种因素控制,变化复杂,加之树体高大,传统测定树木蒸腾量的方法受到很大限制; 而以热脉冲、热扩散和热平衡为主的热技术是国内外用来测定林木蒸腾的重要方法,可更精确地确定树木蒸腾耗水量。近年来,国内学者应用该技术对北方地区树种杨树(Populus deltoides ‘Lux’ex. I-69/55) (刘奉觉等,1993)、棘皮桦(Betula dahurica)和色木槭(Acer mono) (李海涛等,1998)、核桃楸(Juglans mandshurica) (严昌荣等,1999)、油松(Pinus tabulaeformis) (孙鹏森等,2000; 马履一等,2002; 王华田等,2002; 聂立水等,2005; 林平等,2006)、栓皮栎(Quercus variabilis) (王华田等,2002; 聂立水等,2005)、侧柏(Platycladus orientalis) (王华田等,2006)、白桦(Betula platyphylla) (孙慧珍等,2002)、华北落叶松(Larix principis-rupprechtii) (熊伟等,2003)、胡杨(Populus euphratica)和柽柳(Tamarix chinensis) (张小由等,2003)、沙枣(Elaeagnus angustifolia) (张小由等,2006)、红松(Pinus koraiensis) (孙龙等,2007)、新疆杨(Populus alba var. pyramdalis) (桑玉强等,2009)、二白杨(Populus gansuensis) (常学向等,2004)和樟子松(Pinus sylvestris var. mongolica) (吴丽萍等,2003)等,以及降水较为充足的南方地区树种冷杉(Abies fabri) (吴永波等,2005)、湿地松(Pinus elliottii) (李海涛等,2006)、尖峰栲(Castanopsis jianfengensis) (张刚华等,2007)、马占相思(Acacia mangium) (马玲等,2005; 王华等,2008)、青冈栎(Quercus glauca) (张中峰等,2008)、丝栗栲(Castanopsis fargesii) (王兵等,2009)和杉木(Cunninghamia lanceolata) (赵仲辉等,2009)等的树干液流进行了研究,并分析了液流与环境因子的关系。热平衡包裹式植物茎流计已成功用于草本植物液流观测(Baker et al., 1987; Steinberg et al., 1990; Takagi et al., 2006)。毛竹(Phyllostachys edulis)是我国亚热带地区主要优势树种和主要植被恢复树种之一,利用热技术研究其树干液流的报道(侯小金等,2010)较少。一方面由于侯小金等(2010)的研究未能完全揭示毛竹树干液流的季节变化规律,另一方面,同一树种在不同地区的蒸腾差异较大,而且与气象因子的相关性也不同。因此,本文采用美国Dynamax公司生产的包裹式植物茎流计,以野外调查和连续定位观测相结合的方法,并结合气象因子的同步观测,开展毛竹的液流速率及其与环境因子关系研究,以期掌握毛竹的耗水规律以及主导的影响因子,旨在揭示亚热带地区毛竹水循环机制,为毛竹林水资源有效利用提供科学依据。
1 研究地概况研究地位于江西大岗山森林生态站(114°30'— 114°45'E,27°30'—27°50'N)。气候属亚热带季风湿润类型,年均气温15.8~17.7 ℃,月平均最低温度在1月(-5.3 ℃),最高在7月(28.8 ℃),年均降水量为1 591 mm,主要集中在4—6月,年均蒸发量为1 503 mm。天然常绿阔叶林是中亚热带的地带性植被,也是本地区天然林演替系列的顶极群落。由于长期严重的人为干扰,地带性植被已破坏殆尽,现有的各种植被类型主要有天然次生常绿阔叶林、落叶阔叶林、各类针阔混交林、毛竹林以及大面积杉木人工林。本研究试验样地林相结构较整齐,只有极少量马尾松(Pinus massoniana)侵入,海拔315 m,黄壤,东北坡,郁闭度0.7,立竹度2 400株·hm-2。林下植被极少,主要有狗脊(Woodwardia japonica)、苦槠(Castanopsis sclerophylla)、山麦冬(Liriope spicata)、山姜(Alpinia japonica)等。
2 研究方法 2.1 观测样木的确定在具代表性地段设置20 m × 30 m的样方,对毛竹林的胸径和年龄分布进行调查。根据调查结果,以径阶株数和分布范围为观测样木的选择依据,选择径阶8~10 cm、生长良好、树干通直、无被挤压现象的标准木3株,其中每一株分别代表 1度竹、2度竹和3度竹。毛竹新竹长成后,第2年春季换叶,以后每2年换叶1次,每换1次叶竹龄增加1度。根据毛竹生长的特性,通常春季3—5月为生长初期; 6—8月为生长中期; 9—11月,生理活动逐渐缓慢,为生长末期; 12月至翌年2月,气温较低,生理活动最弱,为非生长期。
2.2 树干液流测定 2.2.1 FLOW32包裹式植物茎流计原理Dynamax公司生产的茎干液流测量计用来监测直径在2~150 mm植物的液流速率。茎流计热源以恒定的功率(Pin)作用于茎干后,在不考虑茎干本身热容量的情况下可被分解为3个部分:一部分与垂直方向上的水流进行热交换(Qv); 另一部分以辐射的形式向四周散发(Qr); 还有一部分则与树干内的水流一起向上传输(Qf)。能量平衡方程见文献(严昌荣等,1999; Takagi et al., 2006)。在能量平衡方程中,木本、禾本、空心植物类型对应的轴向热传导常数Kst分别为0.42,0.54,0.28 W·m-1 K-1。毛竹属于禾本科(Gramineae)竹亚科(Bambusoideae)植物,且属空心类植物,因此Kst取0.28。以3株样木液流的平均值作为最终分析值。单木平均液流速率Q(g·h-1) : 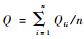
|
|

茎流仪的安装与数据采集:被测样木在测定前,用砂纸轻轻将茎干打磨光滑,然后用游标卡尺精确测定树干直径,之后在打磨好的安装区抹上G4油脂,仔细将包埋式探头安装于被测区,用铝箔包裹,以防止太阳辐射,最后用胶带密封,防止雨水进入。将探头进行数据传输的电缆线与数据采集器相应的接口连接,数据采集器的电源线与12 V电池接通。通过计算机与数据采集器连接,将测定样木的茎干类型、面积、探头电压、起始时间、数据记录的间隔等参数输入到数据采集器,然后定期用电脑采集数据。本研究数据扫描间隔为10 s,每60 min进行平均值计算并储存。2009年9月—2010年7月,采用Dynamax公司生产的FLOW32包裹式茎流计对毛竹树干液流进行了每月连续20~30天的定位观测,7月因为停电等因素,只观测了14天。
2.3 环境因子测定光合有效辐射、总辐射、空气温度、空气湿度、风向等环境因子用HOBO自动气象站测定,数据采集时间与液流仪同步。空气水汽压亏缺(VPD)计算方法见王华等(2008)。
2.4 数据处理及计算运用Excel和SPSS分析软件对数据进行分析和处理。
3 结果与分析 3.1 不同季节不同天气毛竹液流速率日变化不同季节不同天气(晴天和阴天)毛竹液流速率的平均日变化呈现明显的昼夜变化规律(图 1),白天高,夜晚低,而且晴天液流速率的峰值明显高于阴天。晴天,冬、春、夏、秋季液流速率分别在8: 00,6: 00,6: 00和7: 00左右开始稳步升高,随着太阳辐射的逐渐增加,气温增高,空气相对湿度下降,气孔导度不断升高,液流速率逐渐增加,峰值分别出现在14: 00 (180.736 g·h-1),13: 00 (1 508.499 g·h-1),12: 00 (2 737.835 g·h-1)和12: 00 (891.551 g·h-1)前后; 然后随着太阳辐射的减弱,温度降低,叶内外水汽压差减小,液流速率开始逐渐下降,分别在19: 00,21: 00,22: 00和20: 00前后回到低值,至次日4: 00—5: 00液流速率一直维持在较低水平。阴天与晴天类似,只是液流速率开始升高和峰值出现的时刻较晚,回到低值的时刻较早,峰值大为降低; 冬、春、夏、秋季液流速率分别在9: 00,7: 00,7: 00和8: 00左右开始升高,峰值分别出现在14: 00 (108.198 g·h-1),14: 00 (583.078 g·h-1),13: 00 (1 304.763 g·h-1)和13: 00 (354.042 g·h-1)前后,又分别在18: 00,20: 00,21: 00和19: 00前后回到低值,至次日4: 00—5: 00液流速率一直维持在较低水平。白天气温高,光照充足,辐射强,树木蒸腾量大,树干液流速率高; 夜间气温低,树木蒸腾量小,水分通过根压主动吸收进入植物体内,树干液流速率低。由冬季至夏季,光照时间逐渐延长,液流速率从开始升高到回到低值之间的时间逐渐增加; 由夏季至冬季,光照时间逐渐缩短,液流速率从开始升高到回到低值之间的时间逐渐减少。

|
图 1 不同季节不同天气毛竹液流与太阳辐射日变化 Figure 1 Daily changes in sap flow of Phyllostachys edulis and solar radiation in different seasons and weathers |
从图 2、图 3液流和气象因子连日变化曲线可以看出:春、夏、秋、冬各季晴天,辐射、温度和空气水汽压亏缺高,液流速率峰值高,阴雨天气,辐射、温度和空气水汽压亏缺低,液流速率峰值降低; 春、夏、秋、冬四季晴天液流峰值分别在618.14~1 491.96 g·h-1,2 159.44~2 704.46 g·h-1,440.90~681.01 g·h-1和162.85~246.72 g·h-1之间,阴雨天液流峰值分别在70.44~216.04 g·h-1,134.05~140.07 g·h-1,261.82~378.05 g·h-1和29.40~108.20g·h-1之间。秋、冬季毛竹液流速率连日变化曲线波动较小,春、夏季液流速率变化波动较大,这主要是由于研究期间春、夏季降雨较多,而秋、冬季降雨较少所致; 不同季节晴天,毛竹液流速率均呈明显的单峰曲线,春、夏两季受天气因素的影响有时出现双峰曲线,说明毛竹树干液流的“午休”现象并不明显; 不同季节不同天气条件,毛竹夜间的液流微弱,且时断时续,冬季夜间液流速率维持在0.58~10.56 g·h-1之间,春季夜间维持在2.91~38.28 g·h-1之间,而夏季和秋季夜间分别维持在11.16~99.47 g·h-1和1.18~12.39 g·h-1之间; 另外不同季节雨天日间和夜间液流速率没有大的变化,可能是由于雨天,空气湿度大大增加,大气水势也降低,大气与叶之间水势差小。

|
图 2 不同季节毛竹液流连日变化 Figure 2 Sap flow changes of Phyllostachys edulis in consecutive days of different seasons |

|
图 3 不同季节气象因子的连日变化 Figure 3 Changes of the meteorological factors in consecutive days of different seasons |
为了进一步分析各气象因子对毛竹液流速率的影响,通过对1月、4月、7月和10月份树干液流连日变化与气象因子相关分析表明,树干液流速率与光合有效辐射、太阳总辐射、空气温度、风速和VPD均有很强的相关性。1月份表现为空气温度(0.757)>太阳总辐射(0.747)>光合有效辐射(0.744)>VPD(0.726)>风速(0.613); 4月表现为光合有效辐射(0.894)>太阳总辐射(0.893)>VPD(0.673)>风速(0.625)>空气温度(0.619); 7月份表现为光合有效辐射(0.905)>太阳总辐射(0.896)>VPD (0.857)>空气温度(0.842)>风速(0.761); 10月表现为光合有效辐射(0.782)>VPD(0.706)>太阳总辐射(0.624)>空气温度(0.623)>风速(0.457)。不同时期树干液流速率与气象因子的相关性强弱有所差异,非生长季毛竹树干液流速率与空气温度的相关性最强,而生长季毛竹树干液流速率均与光合有效辐射的相关性最强。非生长季毛竹光合速率和蒸腾速率低,主要靠根压以主动吸水方式从土壤中吸水,温度对根压有重要影响,说明空气温度是非生长季毛竹蒸腾作用的最主要影响因子。生长季毛竹光合速率和蒸腾速率高,主要靠叶片蒸腾作用产生的蒸腾拉力进行被动吸水,光合有效辐射是光合作用的驱动力,影响气孔的开张程度,从而影响蒸腾作用,说明光合有效辐射是生长季毛竹蒸腾作用的最主要影响因子。不同时期树干液流速率与风速的相关性均较弱,说明风速对毛竹蒸腾速率的影响较小。
3.3 毛竹液流速率的季节变化从图 4可以看出:毛竹日均液流速率和日均光合有效辐射、太阳辐射、空气温度等均呈现明显的季节变化规律,生长初、中期3—7月的日均液流速率波动大,非生长期12月至次年2月变化较为平缓,而生长末期9—11月波动较小。分析发现,毛竹日均液流速率季节变化差异显著(P<0.000 1)。毛竹一般在3月开始发笋,随着辐射增加,气温回升,毛竹也开始进入生长季节,需要吸收水分来完成体内的各种新陈代谢。从3月中旬开始,毛竹的液流速率开始上升,3月底到4月下旬,由于此间降雨频繁,辐射强度、大气温度以及饱和气压差都明显降低,液流速率下降。从4月下旬到5月液流速率有了较大幅度的提高,到6月又出现明显的频繁波动,这是由于此期间降雨频繁,辐射强度、大气温度以及饱和气压差都明显降低,蒸腾下降。到7月,土壤供水充足,温度升高,辐射增强,气温达到了一年中的最高值,土壤、植物、大气之间形成了足够大的水势差,因此毛竹耗水量急剧加大,液流速率达到研究期间的最高值。进入9月、10月和11月,由于毛竹生长缓慢,大气温度开始下降,降雨量减少,毛竹蒸腾耗水量减少,液流速率下降,一直到12月、1月和2月,气温较低,毛竹生理活动最弱,蒸腾作用较低,液流速率达到研究期间的最低值。毛竹生长末期日均液流速率为(97.88 ± 5.73) g·h-1,非生长期日均液流速率为(52.27 ± 3.66) g·h-1,生长初期日均液流速率为(167.81 ± 20.15) g·h-1,生长中期日均液流速率为(414.04 ± 48.62) g·h-1。生长季树木光合作用和蒸腾作用强,主要靠蒸腾拉力吸水,树干液流速率高; 非生长季树木光合作用和蒸腾作用较弱,主要靠根压从土壤中吸水,树干液流速率低。

|
图 4 毛竹日均液流速率与气象因子日平均值的季节变化 Figure 4 Seasonal changes in average daily sap flow rate of Phyllostachys edulis and meteorological factors |
为了阐明气象因子对毛竹液流速率变化的影响,以液流速率为因变量(y),光合有效辐射(x1)、太阳总辐射(x2)、空气温度(x3)、风速(x4)、VPD (x5)为自变量,采用逐步剔除法进行多元回归分析(引入因子P<0.05,剔除因子P<0.10),建立树干液流与环境因子的回归方程。不同时期的液流速率与气象因子回归方程分别为:
生长末期: y =-58.024 + 0.269x1 -0.334x2 + 5.770x3 -14.743x4 + 62.845x5 (F = 167.48,R2 = 0.548)。
非生长期: y =-14.099 + 0.312x2 + 4.538x3 + 76.309x5 (F = 626.35,R2 = 0.739)。
生长初期: y =-73.224 + 0.583x1 + 7.846x3 + 14.857x4 (F = 529.46,R2 = 0.697)。
生长中期: y = 20.288 + 2.773x1 -3.823x2 + 403.942x5 (F = 639.09,R2 = 0.852)。
回归方程和回归系数均达到了极显著水平(P<0.01),说明利用气象因子可以较好地预测毛竹树干液流变化。从入选回归方程的环境因子可以看出,生长季光合有效辐射对毛竹液流速率有较大影响,而非生长季空气温度对毛竹的液流速率有较大影响。
4 结论与讨论不同季节,不论晴天还是阴天毛竹液流速率表现出明显的昼夜变化规律,白天高,夜晚低。由冬季至夏季,液流开始上升的时刻和峰值达到的时刻逐渐提前,回到低值的时刻逐渐推后,由夏季至冬季,液流开始升高的时刻和峰值达到的时刻逐渐推后,回到低值的时刻逐渐提前; 液流的持续时间与昼长变化规律一致(赵仲辉等,2009)。
不同季节晴天毛竹液流速率呈单峰曲线,而春、夏两季有时出现双峰曲线,但“午休”现象不明显,这与陈建华等(2006)和林琼影等(2008)对毛竹光合速率日变化规律的研究结果不一致,而与叶片蒸腾速率日变化规律相似(林琼影等,2008)。这可能是由于毛竹有地上茎和地下茎,地下茎活动较频繁,极易形成根压,加之叶片蒸腾造成的树体水容亏缺,即便部分叶片水分蒸腾减小,液流仍会维持较高水平,难以观测到晴天树干液流的午休现象(孙慧珍等,2002; 张刚华等,2007); 不同季节不同天气条件毛竹夜间均存在微弱的液流,且时断时续,可能是由于树木白天蒸腾消耗过多的水分导致植物体内储水减少,形成植物体内水势梯度和根压的存在并推动树液流动,补充白天过度的水分蒸腾所造成的树干水分减少(李海涛等,1998; 严昌荣等,1999; 孙鹏森等,2000; 熊伟等,2003; 马玲等,2005; 聂立水等,2005; 吴永波等,2005)。
各个时期毛竹树干液流速率与气象因子的相关性强弱有所差异,非生长季树干液流速率与气温的相关性最强,而生长季节与光合有效辐射的相关性最强。说明空气温度是非生长季毛竹蒸腾作用的最主要影响因子,光合有效辐射是生长季毛竹蒸腾作用的最主要影响因子。另外,本研究所得结论与Granier等(1996)、吴永波等(2005)、李海涛等(2006)、张刚华等(2007)、王兵等(2009)和赵仲辉等(2009)在水分充足地区对热带雨林、冷杉、湿地松、尖峰栲、丝栗栲和杉木的研究结论有所差异。这可能是由于毛竹属多年生禾本植物,木质部中有大量的导管和管胞,加之庞大的地下鞭根系统,像一个巨大的蓄水池,不断地从土壤中吸收植物需要的水分,导致与木本植物液流的差异。
研究表明:毛竹受自身发育状况和各种外界因素的影响树干液流速率表现出显著的季节变化规律。毛竹生长末期、非生长期、生长初期和生长中期日均液流速率分别为(97.88 ± 5.73) g·h-1,(52.27 ± 3.66) g·h-1,(167.81 ± 20.15) g·h-1和(414.04 ± 48.62) g·h-1。因此旱季应通过整地增强林地的蓄水保墒能力,以及密度调控等经营措施减少单木和群体耗水量,保证充足的水分供应,有利于毛竹的出笋和生长。
气象因子是影响植物蒸腾作用的重要因素,在自然条件下,各气象因子并不是独立作用,而是综合影响植物蒸腾(桑玉强等,2009)。本研究分析表明:不同季节毛竹树干液流受光合有效辐射、太阳总辐射、空气温度、水蒸气压亏缺、风速等因素的作用大小是不同的,生长季光合有效辐射对毛竹液流速率有较大影响,非生长季空气温度则对毛竹的液流速率影响较大。这可能是非生长季气温低,叶片生理活性低,光合速率和蒸腾速率低,蒸腾拉力很低,植物为了从土壤中吸水,必须要有足够大的根压值,温度对根压起着决定的作用。而生长季气温高,光照充足,叶片生理活性强,光合速率和蒸腾速率高,蒸腾拉力大,光合有效辐射是光合作用的驱动力,影响气孔的开张程度,从而影响到蒸腾作用的强弱,光合有效辐射对蒸腾作用起着决定的作用。Chelcy等(2004)研究认为在土壤含水量相对充足时,树木耗水则与直接作用于蒸腾作用介质(以叶片为主)的气象因子更为相关。本研究地处中亚热带,雨量充沛,年均降雨量达1 591 mm,但降雨多集中在春、夏两季,秋、冬两季多为旱季,有关旱季土壤水势、土壤温度等环境因子对毛竹液流是否存在影响,影响程度如何,有待进一步研究。
常学向, 赵文智. 2004. 荒漠绿洲农田防护树种二白杨生长季节树干液流的变化[J]. 生态学报, 24(7): 1436-1441. |
陈建华, 毛丹, 马宗艳, 等. 2006. 毛竹叶片的生理特性[J]. 中南林学院学报, 26(6): 76-80. |
侯小金, 谢锦忠, 格日勒图, 等. 2010. 毛竹液流特征及其与环境因子的关系[J]. 生态学杂志, 29(7): 1263-1269. |
李海涛, 陈灵芝. 1998. 应用热脉冲技术对棘皮桦和五角枫树干液流的研究[J]. 北京林业大学学报, 20(1): 1-5. |
李海涛, 向乐, 夏军, 等. 2006. 应用热扩散技术对亚热带红壤区湿地松人工林树干边材液流的研究[J]. 林业科学, 42(10): 32-38. |
林平, 李吉跃, 马达. 2006. 北京山区油松林蒸腾耗水特性研究[J]. 北京林业大学学报, 28(supp. 1): 47-50. |
林琼影, 陈建新, 杨淑贞, 等. 2008. 毛竹气体交换特征[J]. 浙江林学院学报, 25(4): 522-526. |
刘奉觉, 郑世锴, 巨关升. 1993. 用热脉冲速度记录仪(HPVR)测定树干液流[J]. 植物生理学通讯, 29(2): 110-115. |
马履一, 王华田. 2002. 油松边材液流时空变化及其影响因子研究[J]. 北京林业大学学报, 24(3): 23-27. |
马玲, 赵平, 饶兴权, 等. 2005. 马占相思树干液流特征及其与环境因子的关系[J]. 生态学报, 25(9): 2145-2151. |
聂立水, 李吉跃, 翟洪波. 2005. 油松、栓皮栎树干液流速率的比较[J]. 生态学报, 25(8): 1934-1940. |
桑玉强, 郭芳, 张劲松, 等. 2009. 毛乌素沙地新疆杨蒸腾变化规律及其影响因素[J]. 林业科学, 45(9): 66-71. DOI:10.11707/j.1001-7488.20090912 |
孙慧珍, 周晓峰, 赵惠勋. 2002. 白桦树干液流的动态研究[J]. 生态学报, 22(9): 1387-1391. |
孙龙, 王传宽, 杨国亭, 等. 2007. 应用热扩散技术对红松人工林树干液流通量的研究[J]. 林业科学, 43(11): 8-14. DOI:10.3321/j.issn:1001-7488.2007.11.002 |
孙鹏森, 马履一, 王小平, 等. 2000. 油松树干液流的时空变异性研究[J]. 北京林业大学学报, 22(5): 1-6. |
王兵, 郭浩. 2009. 影响丝栗栲树干液流速度的环境因子分析[J]. 南京林业大学学报, 33(1): 43-48. |
王华, 赵平, 蔡锡安, 等. 2008. 马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应[J]. 应用生态学报, 19(2): 225-230. |
王华田, 马履一. 2002. 利用热扩式边材液流探针(TDP)测定树木整株蒸腾耗水量的研究[J]. 植物生态学报, 26(6): 661-667. |
王华田, 赵文飞, 马履一. 2006. 侧柏树干边材液流的空间变化规律及其相关因子[J]. 林业科学, 42(7): 21-27. |
吴丽萍, 王学东, 尉全恩, 等. 2003. 樟子松树干液流的时空变异性研究[J]. 水土保持研究, 10(4): 66-68. |
吴永波, 薛建辉. 2005. 岷江流域冷杉树干液流的动态变化规律[J]. 南京林业大学学报, 29(6): 60-63. |
熊伟, 王彦辉, 徐德应. 2003. 宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应[J]. 林业科学, 39(2): 1-7. DOI:10.11707/j.1001-7488.20030201 |
严昌荣, AlecDowney, 韩兴国, 等. 1999. 北京山区落叶阔叶林中核桃楸在生长中期的树干液流研究[J]. 生态学报, 19(6): 793-797. |
张刚华, 陈步锋, 聂洁珠, 等. 2007. 热带山地雨林尖峰栲边材液流及其与环境因子的关系[J]. 应用生态学报, 18(4): 742-748. |
张小由, 龚家栋, 周茂先, 等. 2003. 应用热脉冲技术对胡杨和柽柳树干液流的研究[J]. 冰川冻土, 25(5): 586-590. |
张小由, 康尔泅, 张智慧, 等. 2006. 沙枣树干液流的动态研究[J]. 中国沙漠, 26(1): 146-151. |
张中峰, 黄玉清, 李先琨, 等. 2008. 岩溶区青冈栎树干液流特征及其与环境因子关系[J]. 中国岩溶, 27(3): 228-234. |
赵仲辉, 康文星, 田大伦, 等. 2009. 湖南会同杉木液流变化及其与环境因子的关系[J]. 林业科学, 45(7): 127-132. DOI:10.11707/j.1001-7488.20090721 |
周晓峰. 1999. 中国森林与生态环境[M]. 北京: 中国林业出版社: 95-99.
|
Baker J M, van Bavel C H M. 1987. Measurement of mass flow of water in the stems of herbaceous plants[J]. Plant Cell Environ, 10(9): 777-782. |
Chelcy R F, Carol E G, Robert J M, et al. 2004. Diurnal and seasonal variability in the radial distribution of sap flow: predicting total stem flow in Pinus taeda trees[J]. Tree Physiology, 24(9): 941-950. DOI:10.1093/treephys/24.9.941 |
Granier A, Huc R, Barigah S T. 1996. Transpiration of natural rain forest and its dependence on climatic factors[J]. Agric For Meteorol, 78(1/2): 19-29. |
Steinberg S L, van Bavel C H M, McFarland M J. 1990. Improved sap flow gauge for woody and herbaceous plants[J]. Agron J, 82(4): 851-854. DOI:10.2134/agronj1990.00021962008200040037x |
Takagi K, Harazono Y, Noguchi S, et al. 2006. Evaluation of the transpiration rate of lotus using the stem heat-balance method[J]. Aquatic Botany, 85(2): 129-136. DOI:10.1016/j.aquabot.2006.03.006 |