 2011, Vol. 47
2011, Vol. 47文章信息
- 薛瑶芹, 张文辉, 周建云, 杨保林, 周民建
- Xue Yaoqin, Zhang Wenhui, Zhou Jianyun, Yang Baolin, Zhou Minjian
- 秦岭南坡不同生境条件下栓皮栎伐桩萌芽特性
- Sprouting and Growth Characteristics of Quercus variabilis Stump after Cutting in Different Habitats of the Southern Slope of the Qinling Mountains
- 林业科学, 2011, 47(7): 57-64.
- Scientia Silvae Sinicae, 2011, 47(7): 57-64.
-
文章历史
- 收稿日期:2010-05-11
- 修回日期:2011-01-27
-
作者相关文章



2. 西部环境与生态教育部重点实验室 杨凌 712100
2. Key Laboratory of Environment and Ecology in West China, Ministry of Education Yangling 712100
栎类植物的自然更新幼苗主要有3个来源:实生幼苗、根系萌苗和伐桩萌苗(Gardiner et al., 1997; Atwood et al., 2009)。伐桩萌苗是栎类森林皆伐后的主要更新方式。伐桩萌苗依靠伐桩上的休眠芽或不定芽发育成萌苗, 进而长成植株(荆涛等, 2002)。相对于实生苗, 伐桩萌苗的高生长和冠幅生长均较为迅速(Lockhart et al., 2007), 可以很快占据干扰所形成的林隙, 在较短时间内恢复林地的郁闭度, 恢复林地生态功能, 提高次生林生物多样性(Kammesheidt, 1998; Bond et al., 2001)。
国外学者针对栎属植物的伐桩更新特点进行了一些研究: Gardiner等(1997)和Lockhart等(2007)指出黑栎(Quercus nigra)和樱皮栎(Quercus pagoda)伐桩萌苗的建成和生长取决于伐桩直径、生境中的光照条件和采伐季节等因素, 强度较大的间伐有利于伐桩存活和萌苗生长, 疏伐萌生丛可以提高萌苗的成活率和生长量; Atwood等(2009)跟踪研究2种栎类植物[沼生栎(Quercus prinus)和白栎(Quercus alba) ]的伐桩在3种不同光照条件下的萌芽特点和萌苗生长特点, 指出皆伐处理下伐桩能产生更多有竞争力的萌苗, 而择伐处理下, 较多的保留木会对伐桩萌发产生消极影响。国内学者对辽东栎(Quercus liaotungensis)伐桩萌芽特点也进行一些研究, 指出伐桩萌苗是辽东栎顺利克服更新瓶颈的一种手段, 在种群更新中起一定作用(高贤明等, 2001; 田丽等, 2007); 坡向、坡度和坡位是影响辽东栎伐桩萌苗的主要立地因子, 而萌苗数量随着伐桩直径的增大先增多后减少, 但与伐桩高度关系不大(伊力塔等, 2006; 2007)。国内学者对于栓皮栎(Quercus variabilis)伐桩萌芽特性的研究极少, 仅有吴明作等(2001)在对河南栓皮栎种群生殖生态研究中指出栓皮栎在受到干扰情况下, 其伐桩萌芽能力能得到迅速而充分地发挥。
栓皮栎是我国暖温带落叶阔叶林和亚热带常绿落叶阔叶混交林的重要组成树种, 分布于我国22个省区, 经济和生态价值大, 在林业生产中起重要作用(张文辉等, 2002)。伐桩萌苗是栓皮栎最常见也是最主要的无性繁殖方式。栓皮栎在分布区内主要作为薪炭林和食用菌培育原料, 多年的过度利用导致绝大多数栓皮栎天然林退化为残败次生林, 生态功能和经济效益明显下降(张文辉等, 2002), 而且林中随处可见农户对栓皮栎的不定期反复砍伐所形成的伐桩。从野外观察到的情况看, 伐桩直径不同, 其上的萌苗数量和萌苗生长量也不同, 有些伐桩在砍伐后几年内就出现衰老特征并部分腐烂。虽然栓皮栎伐桩萌苗在林地恢复中发挥了重要作用, 但伐桩直径和高度对萌苗数量、生长是否有影响, 萌苗数量与萌苗生长的关系如何, 伐桩的萌发能力在砍伐多久就开始下降, 如何利用这些关系进行栓皮栎次生林改造才能使林地的覆盖度增加等等, 对这些问题的研究却未见报道。因此, 本研究以秦岭南坡同一时间砍伐但位于不同坡向的栓皮栎伐桩为研究对象, 研究环境条件、伐桩高度、伐桩直径对萌芽更新的影响, 为栓皮栎次生林改造及种群恢复提供理论依据和技术指导。
1 材料与方法 1.1 研究地区及研究对象研究区域位于秦岭南坡中段, 陕西省佛坪县西北部(107°40'—107°55' E, 33°33'—33°46' N)。该区处于亚热带向暖温带湿润季风气候过渡区, 基带属暖温带气候, 年平均气温6 ~ 14 ℃, 无霜期220天左右, 降水量940 mm (刘新玉等, 2008; 吴燕等, 2008)。地带性植被为暖温带落叶阔叶林, 栓皮栎在本区域形成大面积纯林或混交林。一般林地中, 栓皮栎占到60%以上, 混生树种主要为麻栎(Quercus acutissima)。该区的栓皮栎林一直作为薪炭林或菌用林经营, 由于多年砍伐利用, 目前的栓皮栎林绝大多数沦为残败次生林。
本研究的对象是20世纪70年代以来保护较好的以实生起源为主的栓皮栎纯林(栓皮栎占80%以上)。2007年林权变更后, 部分农户为了获取培养食用菌所需的木料, 对部分林地进行了小面积皆伐, 形成了斑块状的皆伐迹地。迹地上的栓皮栎伐桩高度参差不齐(最高达到70 cm), 伐桩直径也大小不一, 为分析不同生境下伐桩萌生苗生长发育特点及其影响因素提供了良好基础。
1.2 研究方法 1.2.1 野外调查2008年9月, 对2块面积较大的皆伐迹地(伐桩均为2007年冬季砍伐形成)进行调查, 调查内容包括: 1)生境条件:每一块皆伐迹地所处的海拔、坡向、坡度和坡位, 具体见表 1; 2)对伐桩进行标记, 并测量伐桩直径(基径)、高度和伐桩上萌苗数量; 3)萌苗生长特征:分别测量伐桩上每株萌苗的高度、基径和冠幅, 并记录萌苗的年龄。2009年9月进行复查。
|
|
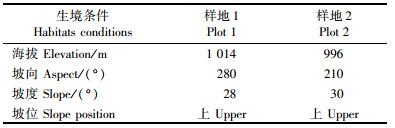
生境划分:本研究所调查的样地, 其海拔、坡度和坡位均相差不多, 由坡向不同而导致的不同水热条件可能对伐桩萌苗建成和生长造成影响, 因此依据坡向将伐桩的生境划分为阳坡和阴坡, 其中阳坡伐桩共122个, 阴坡伐桩共70个。
伐桩径级划分:本研究所调查的伐桩直径在3 ~ 47 cm范围内, 根据伐桩数量在各直径上的分布, 以5 cm为径级区分单位, 共分为7级:伐桩直径≤5 cm为1级, 5 cm<伐桩直径≤ 10 cm为2级, 10 cm<伐桩直径≤15 cm为3级, 依此类推, 伐桩直径>30 cm为7级(李景文等, 2005)。分别统计2个生境下各径级伐桩上的平均萌苗数量及平均萌苗基径、高度和冠幅, 分析伐桩直径与萌苗数量及萌苗生长的关系。
伐桩高度级划分:根据伐桩高度范围(8 ~ 70 cm)以及各高度上伐桩数量, 以10 cm为单位区分高度级, 共分为7级:伐桩高度≤10 cm为1级, 10 cm<伐桩高度≤20 cm为2级, 20<伐桩高度≤30 cm为3级, 依此类推, 高度>60 cm为7级。分别统计2个生境下各高度级伐桩上的平均萌苗数量及平均萌苗基径、高度和冠幅, 分析萌苗数量和生长与伐桩高度的关系。
回归分析:应用SPSS13.0对萌苗基径、高度、冠幅与萌苗数量的关系分别进行回归分析, 采用拟合最好的方程并检验其显著性(赵睿等, 2009)。
综合2008和2009年的调查数据, 计算不同生境中产生当年生萌苗的伐桩占调查伐桩总量的百分比, 即伐桩萌发率。以2008和2009年各伐桩上当年生的萌苗为对象, 统计不同生境下萌苗的平均基径、高度、冠幅。
应用SPSS13.0对统计数据进行单因素方差分析(One-way ANOVA)或多因素方差分析(MultiANOVA) (Johansson, 2008), 并在α = 0.05和α = 0.01水平上检验差异的显著性。
2 结果与分析 2.1 伐桩直径与萌苗生长的关系2个生境下各个径级的伐桩都有萌芽能力(阴坡没有直径<5 cm的伐桩)。多因素方差分析表明:生境条件和伐桩直径对萌苗数量有极显著交互作用(F = 3.545, P<0.01)。总体上看, 阳坡各径级伐桩上的萌苗数量均多于阴坡, 除4和5径级外, 其他径级上的萌苗数量在2个生境间有显著差异(P<0.05) (图 1A)。阴坡的萌苗数量随着伐桩直径增大而增加, 达到一定伐桩直径(20 ~ 25 cm)后又随着伐桩增大而减少; 阳坡萌苗数量变动较大, 与阴坡不同的是, 5径级(直径为20 ~ 25 cm)上的萌苗最少, 平均每桩仅有6个萌苗, 而6和7径级伐桩的萌苗数量可达20个以上(图 1A)。

|
图 1 不同生境下伐桩直径与萌苗数量、萌苗基径、高度及冠幅的关系 Figure 1 Sprouts per stump, sprout base diameter, height and crown width of different stump diameter class in two habitats 数据为平均值±标准误。不同字母代表数据在生境间差异显著(P<0. 05)。Bars are mean value ± standard error. Different letters indicate significant difference among habitats.下同The same below. |
萌苗基径未受到生境条件和伐桩直径交互作用的影响(F = 1.003, P>0.05)。在各径级伐桩上, 阴坡的萌苗基径均大于阳坡, 但大部分径级间没有显著差异(P>0.05) (图 1B), 可能是因为阴坡伐桩上的萌苗数量较少, 同一伐桩上的竞争较为缓和, 因此, 萌苗能获得较多营养物质和生长空间。在阴坡, 随着伐桩增大萌苗基径也逐渐增加, 且单因素方差分析表明萌苗基径在各径级间有显著差异(P<0.05);阳坡的萌苗基径随着伐桩增大而减少, 一直到4径级后, 萌苗的基径才逐渐增加(图 1B), 这可能与同一伐桩上萌苗对有限营养的竞争有关。
生境条件和伐桩直径对萌苗高度没有显著交互作用(F = 0.848, P>0.05), 但2个生境的萌苗高度差异极显著(F = 7.781, P<0.01), 阴坡萌苗高度大于阳坡。在阴坡, 各伐桩径级上萌苗高度均能达到2 m以上, 最大值(2.78 m)出现在5径级, 之后随伐桩增大萌苗高度有所下降; 在阳坡, 萌苗高度先降低后增加, 与萌苗基径的趋势一致, 而且萌苗高度在各径级间有显著差异(P<0.05), 4径级上的萌苗高度最低(1.45 m) (图 1C)。
生境条件和伐桩直径对萌苗冠幅没有显著交互作用(F = 0.848, P>0.05)。除5, 6径级外, 其他径级伐桩上的萌苗冠幅均表现为阳坡大于阴坡, 但差异并未达到显著水平(P>0.05)。萌苗冠幅在2个生境中表现出较为一致的趋势, 即在各径级间有波动, 最小值均出现在5径级(分别为0.9, 0.8 m), 最大值为7径级(1.2, 1.3 m) (图 1D)。在阴坡, 萌苗冠幅在各径级间没有显著差异(P>0.05);阳坡的萌苗冠幅则差异显著。
2.2 伐桩高度与萌苗生长的关系多因素方差分析表明:生境条件和伐桩高度对萌苗数量没有明显交互作用(F = 1.220, P>0.05), 但萌苗数量在不同生境间差异极显著(F = 19.588, P<0.01)。总体而言, 阳坡各高度级上的萌苗数量均多于阴坡(图 2A)。阴坡的萌苗数量随着伐桩高度增加而增多, 到6高度级(50.1 ~ 60 cm)时萌苗数量有所下降(每桩5.0个); 阳坡每桩萌苗数量在各高度级之间有较大波动(图 2A)。单因素方差分析表明:无论是在阳坡还是在阴坡, 萌苗数量在各高度级之间均没有显著差异(P>0.05)。

|
图 2 不同生境下伐桩高度与萌苗数量、萌苗基径、高度及冠幅的关系 Figure 2 Sprouts per stump, sprouts base diameter, height and crown width of different stump height class in two habitats |
伐桩高度和生境对萌苗基径没有显著交互作用(F = 1.381, P>0.05), 但2个生境间的萌苗基径差异极为显著(F = 32.417, P<0.01)。在同一高度级上, 阴坡萌苗基径均大于阳坡(图 2B)。在2个生境中, 随着伐桩高度增加, 萌苗基径均缓慢增加。阳坡的萌苗基径在各伐桩高度级间有显著差异(P<0.05), 而阴坡则无显著差异(P>0.05)。
生境条件和伐桩高度对萌苗高度没有显著交互作用(F = 0.761, P>0.05)。萌苗高度在2个生境中均随着伐桩高度增加而缓慢增加, 除7高度级外, 在同一高度级上的萌苗高度为阴坡显著大于阳坡(P<0.05) (图 2C)。阳坡的萌苗高度在伐桩高度级间有显著差异(P<0.05), 阴坡则无显著差异(P>0.05)。
萌苗冠幅没有受到生境和伐桩高度交互作用的影响(F = 0.344, P>0.05)。在2个生境中, 萌苗冠幅随着伐桩高度增加而呈现缓慢增加的趋势, 各高度级间的萌苗冠幅没有显著差异(P>0.05) (图 2D)。各高度级上的萌苗冠幅基本表现为阳坡大于阴坡, 但差异并不显著(P>0.05) (图 2D)。
2.3 伐桩萌苗数量与萌苗生长的关系对阴坡和阳坡的每桩萌苗数量与同一伐桩上萌苗的平均基径、高度、冠幅的回归分析表明(图 3) :在2个生境中, 萌苗基径与萌苗数量均为显著负相关(P = 0.032, P = 0.000) (图 3A1, B1), 说明萌苗数量对萌苗基径的影响较大; 萌苗高度在2个生境中均呈现出先降低后增加的趋势(图 3A2, B2);萌苗冠幅与萌苗数量的关系在阴坡和阳坡有所不同, 在阳坡, 萌苗冠幅随着每桩萌苗数量增多先增大, 之后保持在一定水平上(图 3B3); 在阴坡, 萌苗冠幅随着萌苗数量增多而逐渐减少(图 3A3)。总体而言, 在阳坡, 萌苗的生长状况与萌苗数量显著相关; 但阴坡则不存在这种现象, 说明阳坡的萌苗竞争更加激烈。

|
图 3 伐桩萌苗数量与萌苗基径、高度、冠幅回归分析 Figure 3 Regression analysis between number of stump sprouts and base diameter, height, crown width A1 ~ A3为阴坡数据,B1 ~ B3为阳坡数据。 A1 ~ A3 represent analysis based on shady slope data, B1 ~ B3 represent analysis based on sunny slope data. |
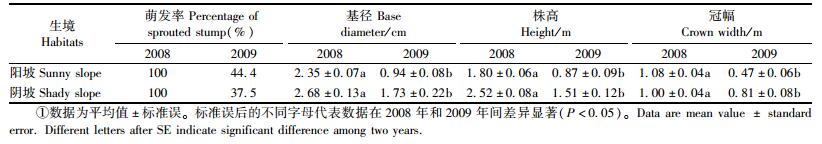
相比于2008年, 2009年的伐桩萌发率在2个生境中均有所下降(分别为44.4%和37.5%) (表 2), 即超过一半的伐桩在2009年没有产生当年生萌苗, 伐桩萌发能力大幅下降, 而且阴坡下降幅度大于阳坡(分别为63.5%和55.6%)。2个生境中当年生萌苗的生长量均呈下降趋势, 而且在2009年当年生萌苗的基径、株高和冠幅均显著低于2008年(P<0.05) (表 2), 萌苗的平均基径和平均高度在阳坡的下降幅度甚至超过50% (分别为60.0%, 51.7%)。另外, 无论是在2008年还是在2009年, 阴坡萌苗的基径、株高和冠幅均大于阳坡, 生长量在2年间的下降幅度也小于阳坡, 2009年阴坡当年生萌苗的基径、高度、冠幅分别下降35.4%, 40.1%和19.0%, 而阳坡萌苗则下降60%, 51.7%和56.4%。这些均表明:砍伐2年后, 伐桩的萌发能力和萌苗生长量均下降, 阴坡伐桩的萌发能力下降幅度较大, 但阳坡萌苗的生长量下降幅度较大。
|
|
萌苗数量和萌苗生长受到环境条件尤其是光照条件的影响。阳坡伐桩上的萌苗数量多于阴坡, 阴坡的萌苗生长优于阳坡; 阳坡萌苗数量与萌苗生长量的关系比阴坡密切。
伐桩直径对萌苗数量和萌苗生长均有影响, 萌苗数量随伐桩直径增大先增加后减少, 直径在15 ~ 25 cm间的伐桩萌苗数量最多, 萌苗生长量随伐桩直径增加先减小后增加; 伐桩高度与萌苗数量的关系不明显, 萌苗生长量随着伐桩高度增加而增加。
砍伐2年后, 伐桩的萌发能力和萌苗生长量均大幅下降, 伐桩内和根系内所贮藏的养分已不能满足萌苗建成和生长的需要。
在栓皮栎次生林改造和经营中, 在水分条件充足的条件下, 应该对基径在15 ~ 25 cm间的栓皮栎个体进行择伐利用, 并且应保留伐桩高度>30 cm, 有利于林地的快速恢复和保持栓皮栎种群规模。在随后的经营利用中, 应疏伐过密的萌苗丛, 控制伐桩的萌发以减缓萌苗间的竞争, 有利于萌苗生长量增加。
4 讨论 4.1 生境条件与伐桩萌苗的关系对比吴明作等(2001)对于河南栓皮栎伐桩萌苗的研究结果, 发现佛坪2个生境的萌苗生长量在砍伐后第1年(2008年)均较大, 萌苗的平均高度分别为1.80和2.52 m, 而河南2, 3年生的栓皮栎萌苗的高度只有0.8 ~ 1.0 m, 这可能是因为本研究选择的区域位于秦岭南坡, 水热条件较好, 而且所有调查的伐桩均是在非生长季(2007年冬季)砍伐, 采伐截断了树根吸收的养分和水分的向上运输, 导致地下部分保存了大量的养分和水分, 而萌苗的生长又恰好处在整个生长季节, 因而有较大的生长量(荆涛等, 2002)。Johansson等(2008)也指出生长季前或者生长季后进行采伐有利于增加桦类(Betula)植物萌苗的生物量。阳坡伐桩上的萌苗数量多于阴坡, 伐桩萌发率也高于阴坡, 这说明环境条件, 尤其是光照条件对伐桩的萌苗数量有较大影响, 该结论与Gardiner等(1997)、Lockhart等(2007)和Atwood等(2009)对不同光照条件下黑栎、樱皮栎、沼生栎和白栎的伐桩萌发特性研究一致, 即光照充足的环境有利于伐桩萌苗的产生。本研究发现生境条件对萌苗的生长有影响, 阴坡的萌苗基径和高度大于阳坡, 而且阴、阳坡不同的环境条件也导致萌苗数量与萌苗基径、高度、冠幅的关系有所不同, 阳坡的萌苗间竞争更为激烈, 导致萌苗数量与萌苗生长的关系较阴坡密切。
4.2 伐桩直径与伐桩萌苗的关系栓皮栎萌苗数量随着伐桩直径的增大先增加后减少, 这与水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、黄檗(Phellodendron amurense)、绒毛桦(Betula pubescens)和欧洲白桦(Betula pendula)等树种的伐桩萌苗规律一致(李景文等, 2005; Lockhart et al., 2007; Johansson et al., 2008)。这主要是因为伐桩萌芽的物质基础是休眠芽和不定芽, 而由原生分生组织形成的休眠芽数量有限, 随着树木的生长其数量逐渐减少(李景文等, 2000), 并随着伐桩直径增大, 树皮变得更加坚硬, 休眠芽冲破树皮的阻力也逐渐增加(Lockhart et al., 2007); 由次生分生组织所产生的不定芽却随着树木主干直径的增大而增多, 但增长到一定程度后, 伐桩产生不定芽的能力与机会越来越少(李景文等, 2000)。萌苗的生长量随着伐桩直径增加先减小后增加, 主要是因为伐桩上的萌苗生长与被砍伐前个体的生长状况也有密切关系(Johansson, 2008), 直径较小的伐桩其伐前个体年龄较小、根系发育不完全、提供的养分和水分有限, 而且萌苗数量逐渐增多导致的竞争也限制萌苗的生长, 4 ~ 5径级的伐桩在采伐前根系发育完全, 伐桩体内贮藏的营养物质增多, 能完全满足休眠芽的活动和萌苗的生长。
4.3 伐桩高度与伐桩萌苗的关系本研究中伐桩高度与萌苗数量的关系并不明显, 这与伊力塔等(2007)对辽东栎以及Mostacedo等(2009)对Centrolodbium microchaete的萌苗数量与伐桩高度关系研究结论相同; 但也有结论相反的研究(荆涛等, 2002; 李景文等, 2005), 这可能是因为栓皮栎的萌苗不仅能出现在伐桩切口周围, 而且伐桩基部地表以下的不定芽也能萌发, 而水曲柳的萌条则主要出现在伐桩切口周围(荆涛等, 2002)。伐桩高度对萌苗的生长有较大影响, 随着伐桩高度增加, 萌苗生长量也有所增加, 这可能是因为伐桩高度越大, 贮存在伐桩内部的营养物质越多, 能够供应萌苗生长的营养物质也越多。
4.4 萌苗数量与萌苗生长的关系Midgley(1996)曾提出萌条能力-高度权衡假说, 认为萌苗生长需要消耗大量的资源, 产生较多萌苗虽然能够提高更新成功率, 但也可能影响到其他方面, 比如高生长。Vesk等(2004)对不同地区植物群落组成的调查显示, 萌发能力强的物种其潜在高度低于萌发能力弱的物种; 但也有结论相反的研究(Nzunda et al., 2007)。这些研究主要针对不同物种进行比较, 可能因为物种的其他生活史特征的差异降低了结果的可比性(赵睿等, 2009)。赵睿等(2009)对米心水青冈(Fagus engeriana)植株丛的萌苗数量与萌苗高度的关系、有萌苗个体和无萌苗个体的高度所进行的对比研究弥补了这一不足, 且证明米心水青冈的高生长并不符合Midgley的假说, 而是随着萌苗数量增加, 萌苗高度也逐渐增加。本研究中栓皮栎的高生长与该假说的观点相反, 与赵睿等(2009)的结论相同; 而萌苗基径随着萌苗数量增多而减小, 这说明砍伐后第1年, 栓皮栎伐桩的萌苗数量实际上对萌苗的基径生长影响更大。随着砍伐时间的增长, 这种关系会不会改变, 还有待于进一步研究。
4.5 伐桩的萌发能力栓皮栎伐桩产生当年生萌苗的能力在砍伐后第2年就开始下降, 而且降幅比较大, 能产生当年生萌苗的伐桩比例不到50%。Imanishi等(2009)在对7个物种砍伐3年后的伐桩萌芽特性研究中也指出:砍伐后第1年大部分物种超过80%的伐桩均能萌发, 而在砍伐后第2年或第3年, 实际萌发的伐桩比例有所下降, 而且萌苗数量也有一定程度下降。樱皮栎伐桩的萌发能力在砍伐后7年中也持续下降(Lockhart et al., 2007)。伐桩萌苗产生是一个大量消耗资源的过程, 有研究指出:有萌发能力的植物, 其根系中的淀粉含量比没有萌发能力的物种高4 ~ 35倍, 碳水化合物含量也较高; 但萌发以后根系中的淀粉含量大幅下降(Bowen et al., 1993; Sakai et al., 1997; Bell et al., 1999)。另外, 贮藏在伐桩和根系中的营养物质会被逐渐消耗以供给萌苗生长, 而不成熟的萌苗不能制造足够的光合产物以弥补这种损耗, 再加上由原生分生组织形成的休眠芽数量有限, 因此只有那些贮存资源多而且有丰富的休眠芽的伐桩才能持续产生萌苗; 但萌发量是否会持续下降, 还是会随着萌苗的逐渐成熟能够提供足够的光合产物供给根系而有所改善, 还需要进一步跟踪观测研究。砍伐后第2年, 当年生萌苗的基径、高度和冠幅均远远小于第1年, 而且该类萌苗对空间和资源竞争中均处于劣势, 能否成活是很大的问题。一旦有萌苗枯死, 实际上是对伐桩资源的浪费, 所以应该控制伐桩的萌发数量并继续深入研究每个伐桩上应该保留几个萌苗才能使萌苗的生物量达到最大, 最有利于林地生物量的增加。
高贤明, 王巍, 杜晓军, 等. 2001. 北京山区辽东栎林的径级结构、种群起源及生态学意义[J]. 植物生态学报, 25(6): 673-678. |
荆涛, 马万里, KujansuuJ, 等. 2002. 水曲柳萌芽更新的研究[J]. 北京林业大学学报, 24(4): 12-15. |
李景文, 刘世英, 王清海, 等. 2000. 三江平原低山丘陵区水曲柳无性更新研究[J]. 植物研究, 20(2): 215-220. |
李景文, 聂绍荃, 安滨河. 2005. 东北东部林区次生林主要阔叶树种的萌芽更新规律[J]. 林业科学, 41(6): 72-77. DOI:10.11707/j.1001-7488.20050611 |
刘新玉, 张泽钧, 郑晓燕, 等. 2008. 佛坪自然保护区6种有蹄动物的活动痕迹监测[J]. 生态学报, 28(9): 4582-4588. |
田丽, 王孝安, 郭华, 等. 2007. 黄土高原马栏林区辽东栎更新特性研究[J]. 广西植物, 27(2): 191-196. |
吴明作, 刘玉萃, 姜志林. 2001. 栓皮栎种群生殖生态与稳定性机制研究[J]. 生态学报, 21(2): 225-230. |
吴燕, 何祥博, 刘雪华. 2008. 陕西佛坪自然保护区大熊猫主食竹巴山木竹林间伐后的质量状况分析[J]. 林业调查规划, 33(5): 63-70. |
伊力塔, 韩海荣, 马钦彦, 等. 2006. 灵空山辽东栎萌芽更新的灰色关联分析[J]. 山西林业科技, (1): 23-25. |
伊力塔, 韩海荣. 2007. 山西灵空山林区辽东栎萌芽更新规律研究[J]. 林业资源管理, (4): 57-61. |
张文辉, 卢志军. 2002. 栓皮栎种群的生物学生态学特性和地理分布研究[J]. 西北植物学报, 22(5): 1093-1101. |
赵睿, 周学峰, 徐娜娜, 等. 2009. 米心水青冈种群萌条更新与高度生长[J]. 生态学报, 29(7): 3665-3669. |
Atwood C J, Fox T R, Loftis D L. 2009. Effects of alternative silviculture on stump sprouting in the southern Appalachians[J]. Forest Ecology and Management, 257(4): 1305-1313. DOI:10.1016/j.foreco.2008.11.028 |
Bell T L, Ojeda F. 1999. Underground starch storage in Erica species of the Cape Floristic Region: differences between seeders and resprouters[J]. New Phytol, 144(1): 143-152. DOI:10.1046/j.1469-8137.1999.00489.x |
Bond W J, Midgley J J. 2001. Ecology of sprouting in woody plants: the persistence niche[J]. Trend in Ecology & Evolution, 16(1): 45-51. |
Bowen B J, Pate J S. 1993. The significance of root starch in post-fire shoot recovery of the resprouter Stirlingia latifolia R[J]. Br. (Proteaceae). Annals of Botany, 72(1): 7-16. DOI:10.1006/anbo.1993.1075 |
Gardiner E S, Helmig L. 1997. Development of water oak stump sprouts under a partial overstory[J]. New Forest, 14(1): 55-62. DOI:10.1023/A:1006502107495 |
Imanishi A, Morimoto J, Imanishi J, et al. 2009. Sprout initiation and growth for three years after cutting in an abandoned secondary forest in Kyoto, Japan[J]. Landscape and Ecological Engineering, 6(2): 325-333. |
Johansson T. 2008. Sprouting ability and biomass production of downy and silver birch stumps of different diameters[J]. Biomass and Bioenergy, 32(10): 944-951. DOI:10.1016/j.biombioe.2008.01.009 |
Kammesheidt L. 1998. The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-andburn agriculture in Eastern Paraguay[J]. Plant Ecology, 139(2): 155-165. DOI:10.1023/A:1009763402998 |
Lockhart B R, Chambers J L. 2007. Cherrybark oak stump sprout survival and development five years following plantation thinning in the lower Mississippi alluvial valley, USA[J]. New Forests, 33(2): 183-192. DOI:10.1007/s11056-006-9022-7 |
Midgley J J. 1996. Why the world's vegetation is not totally dominated by resprouting plants; because resprouters are shorter than reseeders[J]. Ecography, 19(1): 92-94. DOI:10.1111/eco.1996.19.issue-1 |
Mostacedo B, Putz F E, Fredericksen T S, et al. 2009. Contributions of root and stump sprouts to natural regeneration of a logged tropical dry forest in Bolivia[J]. Forest Ecology and Management, 258(6): 978-985. DOI:10.1016/j.foreco.2008.09.059 |
Nzunda E F, Griffiths M E, Lawes M J. 2007. Multi-stemmed trees in subtropical coastal dune forest: survival strategy in response to chronic disturbance[J]. Journal of Vegetation Science, 18(5): 693-700. DOI:10.1111/j.1654-1103.2007.tb02583.x |
Sakai A, Sakai S, Akiyama F. 1997. Do sprouting tree species on erosion-prone sites carry large reserves of resources?[J]. Annals of Botany, 79(6): 625-630. DOI:10.1006/anbo.1996.0389 |
Vesk P A, Warton D I, Westoby M. 2004. Sprouting by semi-arid plants: testing a dichotomy and predictive traits[J]. Oikos, 107(1): 72-89. DOI:10.1111/oik.2004.107.issue-1 |