 2011, Vol. 47
2011, Vol. 47文章信息
- 李海奎, 雷渊才, 曾伟生
- Li Haikui, Lei Yuancai, Zeng Weisheng
- 基于森林清查资料的中国森林植被碳储量
- Forest Carbon Storage in China Estimated Using Forestry Inventory Data
- 林业科学, 2011, 47(7): 7-12.
- Scientia Silvae Sinicae, 2011, 47(7): 7-12.
-
文章历史
- 收稿日期:2010-03-31
- 修回日期:2011-06-07
-
作者相关文章


森林是陆地上最大的生态系统, 与陆地其他生态系统相比, 它具有复杂的层次结构、很长的生命周期、最高的生物量和生长量, 是陆地生物光合产物的主体, 也是陆地生态系统的最大碳库, 约80%的地上碳储量和40%的地下碳储量发生在森林生态系统(Kramer, 1981; Cao et al., 1998)。森林极易受到自然及人类活动的干扰, 进而对全球碳循环过程产生重大影响。通过植树造林实现林业碳汇功能是当前最为有效和快捷的碳平衡方法之一。
中国是《京都议定书》等多个国际性公约的签约国, 作为世界上最大的发展中国家, 在环境恶化、气候变暖的背景条件下, 承担着自身发展和维护、改善世界生态环境的双重责任和压力。中国承诺到2020年全国森林面积比2005年增加4 000万hm2, 森林蓄积量比2005年增加13亿m3, 通过大力增加森林碳汇来减缓温室气体排放。中国地域广阔, 跨越温带至热带的各个气候带, 自然条件复杂, 乔木种类繁多, 森林资源丰富, 森林类型多样, 森林资源在世界上占有相当重要的地位, 森林面积和蓄积均居世界前列, 人工林面积位居世界第一。本研究充分利用第七次全国森林资源连续清查数据, 采用加权回归模型估计法或模型法估算了我国乔木林、竹林、灌木林、疏林地、散生木和四旁树的碳储量; 按区域、起源、龄组和优势树种等分类方法对全国乔木林碳储量和碳密度进行评估, 并分析乔木林碳储量计算方法的不确定性, 对于客观反映我国森林对全球碳循环及全球气候变化的影响、加快森林碳循环研究具有重要意义。
1 数据来源本研究数据来源于第七次全国森林资源连续清查的基础资料:全国范围实测数据包括约66万个样地实测数据、约420万个样木实测数据和约15万个样木树高实测数据; 分省统计结果包括乔木林按优势树种各龄组面积、蓄积统计表, 天然、人工乔木林各龄组面积、蓄积统计表, 疏林地按优势树种面积、蓄积统计表, 灌木林按类型面积统计表和竹林按林种面积、株数统计表。
2 研究方法本次计算的森林植被包括乔木林、竹林、疏林地、散生木、四旁树和纯粹的灌木林(不包括林下灌木), 不包括没有计算蓄积的经济林。森林植被的碳储量估算包括地上和地下部分, 不包括土壤中的碳储量。台湾省、香港特别行政区和澳门特别行政区。
2.1 乔木林生物量计算方法生物量经验(回归)模型估计是利用某一树种野外调查的生物量实测数据, 建立生物量与树高、胸径等统计回归关系模型(Montagu et al., 2005; Brandeis et al., 2006; Lehtonen et al., 2004; Vallet et al., 2009; 胥辉等; 2001;曾伟生等, 1999; 张会儒等, 1999), 可以分别建立树干、树枝、树叶和树根生物量模型(Djomoa et al., 2010; Repola, 2008; 2009; 胥辉等, 2002; 曾伟生等, 2011), 也可以建立与材积相容的单木生物量回归模型(Fournier et al., 2003; Brandeis et al., 2006; Nogueira et al., 2008)。
生物量经验(回归)模型估计法在区域(省级)尺度上估算生物量时, 分省某一优势树种的生物量B1的计算公式为
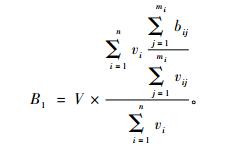
|
式中: V为分省该优势树种的蓄积量; vi为分省该优势树种第i个样地的调查材积; bij为分省该优势树种第i个样地第j个径阶的模型生物量(李海奎等, 2010); vij为分省该优势树种第i个样地第j个径阶的模型材积, 样地模型材积利用二元材积公式v = a + b(D2H)计算(v为模型材积, D为胸径, H为树高, a和b为参数); mi为分省该优势树种第i个样地内径阶数; n为分省该优势树种计算生物量的优势树种样地个数。
样地模型材积和模型生物量计算时, 需要用到各径阶林木树高值, 它的精度对生物量计算结果有很大影响。第七次清查时, 利用全国约15万个树高测定单木样本, 分49个树种, 采用树高分级的方法拟合了树高曲线模型, 模型的形式为Richard方程: H = a(1-e-bD)c, a, b和c为参数, 具体数值见李海奎等(2010)附录2。
鉴于回归模型估计法测算生物量过程比较复杂, 这里列出其详细算法。1)各省树高测定表的处理:统一各省树种代码; 剔出树高小于2 m的样本; 剔出异常样本; 统一单位。2)处理后各省树高测定表合并成全国树高测定表, 按树种、省份和样地号进行排序。3)分树种形成单一树种全国树高表文件。4)分树种确定树高曲线: ①胸径按2 cm整化径阶; ②把树高分为3 ~ 9个不同等级; ③进行树高曲线拟合; ④按照各个样本的树高预测值和实测值的差值, 与哪条曲线最近的原则, 对所有样本再次分组; ⑤反复进行步骤③、④, 直到分组不再变化; ⑥对同一样地多株树木, 按不同等级预测树高与实测树高之差的平方和最小为该样地所有树木的最终分类; ⑦按最终分类, 求解树高曲线各个等级的参数。5)各树种树高表合并成全国树高等级表, 并按省份、树种、样地号进行排序。6)提取各省树高等级表, 并剔出同一样地、同一树种的重复样本。7)样木表的处理:仅保留立木类型为11(有林地)的样木, 去掉检尺类型为14 (已被采伐的林木)、17(错测林木)和18 (生长变化异常的林木)的样木, 并按样地号、树种排序。8)根据各省树高表等级中样地、树种的分级, 对样木表的各个样木进行分级。9)计算样木表中各个样木的树高。10)根据胸径和树高计算各株样木的理论材积(按照相应的公式)。11)根据树种生物量公式计算各株树木的生物量(地上和地下)。12)计算各省分树种的生物量转换系数。13)计算各省分树种的生物量。14)分省合计各树种生物量, 获得省级区域乔木林生物量。
2.2 竹林生物量计算方法单株竹子地上部分生物量Wt计算公式(孙天任, 1986)为Wt = 0. 643 9D1.537 3; 单株竹子地下部分生物量Wr计算公式为Wr = 0. 340 4D1.189 9 +0. 308 7D1.289 2; 分省竹林总生物量B2计算公式为B2 = (Wt + Wr) N。式中: D为分省单株竹子平均胸径; N为总株数。
2.3 灌木林生物量计算方法分省灌木林总生物量B3计算公式为B3 = bA。式中: b为分省灌木林单位面积生物量; A为分省灌木林总面积。
灌木林生物量分省进行, 其中关键的灌木林单位面积生物量(b)根据有关文献(万里强等, 2001; 郑绍伟等, 2007; 胡会峰等, 2006; 黄玫等, 2006; 刘国华等, 2003; 陈遐林等, 2002)和本省乔木林的单位面积生物量综合考虑确定, 由于没有资料可以进行验证, 计算结果精度未知, 但总的计算结果趋于保守。
2.4 疏林、散生木和四旁树生物量计算方法基本公式与乔木林生物量计算公式相同, 分省分树种计算, 合计得到省级区域的疏林、散生木和四旁树生物量, 然后累计所有省份得到全国的疏林、散生木和四旁树总生物量。
2.5 森林碳储量计算方法目前, 无论在森林群落或森林生态系统尺度上还是在区域、国家尺度上, IPCC等国际组织以及世界各国普遍采用的森林碳储量估算方法是直接或间接测定森林生物量, 再乘以生物量中碳元素含量(含碳系数)推算而得。
分省某一优势树种碳储量C计算公式为C = Bc, 式中: B为分省该优势树种生物量; c为该优
中国森林植被碳储量总量为781 146. 08万t, 其中乔木林666 221. 08万t, 占85. 29 %; 疏林地、散生木和四旁树59 281. 71万t, 占7. 59 %; 灌木林35 780. 12万t, 占4. 58%;竹林19 863. 17万t, 占2. 54%。乔木林碳密度为42. 82 t·hm-2。
3.2 中国森林植被碳储量地理分布格局表 2是中国分大区的森林植被碳储量和碳密度表。中国森林植被碳储量主要集中在东北和西南两个大区, 分别占全国的20. 03%和40. 07%, 其乔木林碳密度大于全国平均水平。
|
|

表 3为中国分省森林植被碳储量和碳密度。表 3显示:西藏、黑龙江、云南、四川、内蒙古和吉林6省区森林碳储量最大, 均占全国的5%以上, 6省市合计占全国森林碳储量的60%以上, 而上海、天津、宁夏、北京、青海、江苏、河北、海南、山东和山西10省(市、区)最小、均不到全国的1%, 10省(市、区)合计占全国森林碳储量不足4%。
|
|

西藏、新疆、吉林、海南、四川、云南、青海、黑龙江、福建和甘肃10省(市、区)碳密度大于全国平均水平, 其中西藏最大是全国平均水平的2. 39倍; 小于全国平均水平的21个省(市、区)中, 最小的是上海, 其碳密度不足平均水平的28%。
3.3 中国乔木林分起源、龄组碳储量分布状况由表 4可见:中国乔木林总碳储量为666 221. 08万t, 其中天然林553 285. 91万t, 占83. 05%, 人工林112 935. 17万t, 占16. 95%。在天然林中, 幼龄林、中龄林和近熟林的总碳储量占61. 85%, 成和过熟林占38. 15%;在人工林中, 幼龄林、中龄林和近熟林的总生物占81. 35%, 成熟林和过熟林占18. 65%。
|
|

由表 5可见:天然林各个龄组碳密度均大于人工林, 但在幼龄林和中龄林差别不大, 近熟林以后差别变大, 这可能是由于天然林和人工林龄组分组的不一致造成的, 也间接说明人工林尚有一定的潜力。除了人工林在过熟林时碳密度略低于成熟林外, 碳密度总的趋势都是过熟林>成熟林>近熟林>中龄林>幼龄林。天然林总的碳密度大于乔木林的平均值(42. 82 t·hm-2), 而人工林总的碳密度小于乔木林的平均值, 这说明人工林中幼龄林占多数。
|
|
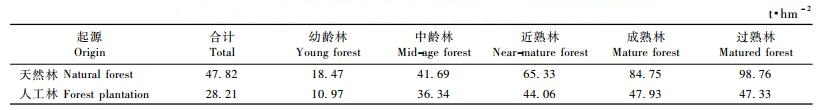
表 6显示:阔叶混、栎(Quercus)类、杉木(Cunninghamia lanceolata)、针阔混、其他硬阔类、杨(Populus)树、落叶松(Larix)和白桦(Betula platyphylla)等树种的碳储量较大, 均占全部碳储量的5%以上, 除去混交树种(组), 只有栎类、杉木、杨树、落叶松和白桦5个树种(组), 这5个树种组占总碳储量的33%;碳储量占总碳储量不到0. 1%的树种(组)有楝树(Melia azedarach)、檫木(Sassafras)、池杉(Taxodiam ascendens)、火炬松(Pinus taeda)、楠木(Phoebe)、木麻黄(Casuarina equisetifolia)、其他松类、泡桐(Paulawnia fortunei)、樟木(Cinnamomum camphora)、水杉(Metasequoia glyptostroboides)、国外松、枫桦(Betula costata)、相思(Acacia)、黑松(Pinus thunbergii)、枫香(Liquidamber)、赤松(Pinus densiflora)和油杉(Keteleeria), 17个树种合计碳储量占总碳储量的比例不到1%。
|
|

碳密度较大的树种(组)有高山松(Pinus densata)、冷杉(Abies)、铁杉(Tsuga)、云杉(Picea)、池杉、阔叶混、樟子松(Pinus sylvestrisa)、思茅松(Pinus kesiya var. langbiamensis)、椴树(Tilia)、水杉(Metasequoid glyptostroboides)、楠木、其他硬阔类、柳杉(Cryptomeria fortunei)、栎类和针阔混, 高山松最大, 是平均值的2. 14倍, 碳密度较小的树种(组)有楝树、其他松类、桉树(Eucalyptus)、榆树(Ulmus)、柳树(Salix)、檫木、湿地松(Pinus elliottii)、桦木(Betula)、油杉等, 其中楝树最小, 不足平均数的20%。
表 7显示:阔叶树总碳储量大于针叶树总碳储量, 其碳密度也较大。
|
|

中国森林植被碳储量总量中乔木林所占比例超过85%, 是主体部分; 中国森林植被碳储量主要集中在西南和东北2大区, 占全国的比例超过60%, 这2个地区乔木林碳密度高于全国平均水平, 和中国森林的分布一致; 中国乔木林碳储量中人工林的比例超过15%, 且人工林中幼龄林、中龄林的比例较多, 可以预测未来人工林在乔木林碳储量中的比例会不断提高; 中国乔木林碳储量主要集中于阔叶混、栎类和杉木、针阔混、其他硬阔类、杨树、落叶松和白桦等树种(组), 而碳密度大于平均值的树种(组)为高山松、冷杉、铁杉、云杉、池杉、阔叶混、樟子松、思茅松、椴树、水杉、楠木、其他硬阔类、柳杉、栎类和针阔混等, 阔叶混、栎类、其他硬阔类和针阔混4个树种(组)的碳储量和碳密度均较高, 但总体来讲, 阔叶树的碳储量和碳密度均大于针叶树。
基于森林资源连续清查数据估算全国乔木林碳储量, 采用生物量经验(回归)模型估计法时, 是分省、分树种计算的, 这时通常会存在3个问题: 1)地方性模型地域性强且覆盖度低, 缺乏经过验证的全国性通用模型; 2)由于范围大、立地条件复杂, 具体表现为同一树种、同一胸径在不同的地区树高差别很大, 所以生物量模型多为胸径和树高的二元模型, 而清查数据的样木通常只有胸径而没有树高; 3)由样木到样地, 再到统计单位(省)的转换系数扩展问题。对第1个问题, 现阶段没有更好的方法, 本研究采用的是已有的研究成果1)和收集的模型, 好在国家林业局已在2009年开始的第八次全国森林资源连续清查中增加了国家层面上的生物量建模外业调查工作并开展了建模方法研究, 相信在八次清查汇总时估算生物量会有较好的适用性模型; 对第2个问题, 本研究采用了第七次清查时的树高测定数据, 全国分为49个树种(组), 由于面积和分布范围差异, 每个树种的样本从最少的32个到最多的19 017个不等, 拟合的树高曲线模型决定系数大多大于0. 9(只有2个树种为0. 85 ~ 0. 9), 均方根误差(RMSE)为0. 56 ~ 2. 00 m (平均高6. 5 ~ 17. 9 m), 均方根相对误差为6. 03% ~ 14. 13%, 其中39个树种小于10%, 基本上满足了精度要求; 对于第3个问题, 本研究采用了由样木加权到径阶、再到样地、最后扩展到统计单位, 使用了样地中所有的样木信息。
1) 唐守正. 1999.二元立木生物量模型及其相容的一元自适应模型系列研究报告
陈遐林, 马钦彦, 康峰峰, 等. 2002. 山西太岳山典型灌木林生物量及生产力研究[J]. 林业科学研究, 15(3): 304-309. |
胡会峰, 王志恒, 刘国华, 等. 2006. 中国主要灌丛植被碳储量[J]. 植物生态学报, 30(4): 539-544. |
黄玫, 季劲钧, 曹明奎, 等. 2006. 中国区域植被地上与地下生物量模拟[J]. 生态学报, 26(12): 4156-4163. DOI:10.3321/j.issn:1000-0933.2006.12.031 |
李海奎, 雷渊才. 2010. 中国森林植被生物量和碳储量评估[M]. 北京: 中国林业出版社.
|
刘国华, 马克明, 傅伯杰, 等. 2003. 岷江干旱河谷主要灌丛类型地上生物量研究[J]. 生态学报, 23(9): 1757-1763. |
孙天任. 1986. 水竹(Phyllostachys heteroclada)人工林生物量结构研究[J]. 植物生态学与地植物学报, 10(3): 190-198. |
万里强, 李向林, 苏加楷, 等. 2001. 长江三峡地区灌木生物量及产量估测模型[J]. 草业科学, 18(5): 5-10. |
胥辉, 刘伟平. 2001. 相容性生物量模型研究[J]. 福建林学院学报, 21(1): 18-23. |
胥辉, 张会儒. 2002. 树木生物量模型研究[M]. 昆明: 云南科技出版社.
|
曾伟生, 骆期邦, 贺东北. 1999. 兼容性立木生物量非线性模型研究[J]. 生态学杂志, 18(4): 19-24. |
曾伟生, 张会儒, 唐守正. 2011. 立木生物量建模方法[M]. 北京: 中国林业出版社.
|
张会儒, 唐守正, 胥辉. 1999. 关于生物量模型中的异方差问题[J]. 林业资源管理, (1): 146-149. |
郑绍伟, 唐敏, 邹俊辉, 等. 2007. 灌木群落及生物量研究综述[J]. 成都大学学报:自然科学版, 26(3): 189-192. |
Brandeis T J, Delaney M, Parresol B R, et al. 2006. Development of equations for predicting Puerto Rican subtropical dry forest biomass and volume[J]. Forest Ecology and Management, 233(1): 133-142. DOI:10.1016/j.foreco.2006.06.012 |
Cao M, Woodward F I. 1998. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their response to climate change[J]. Global Change Biology, 4(2): 185-198. DOI:10.1046/j.1365-2486.1998.00125.x |
Djomoa A N, Ibrahimab A, Saborowskic J, et al. 2010. Allometric equations for biomass estimations in Cameroon and pan moist tropical equations including biomass data from Africa[J]. Forest Ecology and Management, 260(10): 1873-1885. DOI:10.1016/j.foreco.2010.08.034 |
Fournier R A, Luther J E, Guindon L, et al. 2003. Mapping aboveground tree biomass at the stand level from inventory information: test cases in Newfoundland and Quebec[J]. Canadian Journal of Forest Research, 33(10): 1846-1863. DOI:10.1139/x03-099 |
Kramer P J. 1981. Carbon dioxide concentration, photosynthesis, and dry matter production[J]. BioScience, 31(1): 29-33. DOI:10.2307/1308175 |
Lehtonen A, Mäkipää R, Heikkinenb J, et al. 2004. Biomass expansion factors (BEFs) for Scots pine, Norway spruce and birch according to stand age for boreal forests[J]. Forest Ecology and Management, 188(1/3): 211-224. |
Montagu K D, Düttmer K, Bartona C V M, et al. 2005. Developing general allometric relationships for regional estimates of carbon sequestration—an example using Eucalyptus pilularis from seven contrasting sites[J]. Forest Ecology and Management, 204(1): 115-129. DOI:10.1016/j.foreco.2004.09.003 |
Nogueira E M, Fearnside P M, Nelson B W, et al. 2008. Estimates of forest biomass in the Brazilian Amazon: new allometric equations and adjustments to biomass from wood-volume inventories[J]. Forest Ecology and Management, 256(11): 1853-1867. DOI:10.1016/j.foreco.2008.07.022 |
Repola J. 2008. Biomass equations for birch in Finland[J]. Silva Fennica, 42(4): 605-624. |
Repola J. 2009. Biomass equations for Scots pine and Norway spruce in Finland[J]. Silva Fennica, 43(4): 625-647. |
Vallet P, Meredieu C, Seynave I, et al. 2009. Species substitution for carbon storage: sessile oak versus Corsican pine in France as a case study[J]. Forest Ecology and Management, 257(4): 1314-1323. DOI:10.1016/j.foreco.2008.11.034 |