 2011, Vol. 47
2011, Vol. 47文章信息
- 杨桦, 杨伟, 杨茂发, 杨春平, 潘武全, 冯波
- Yang Hua, Yang Wei, Yang Maofa, Yang Chunping, Pan Wuquan, Feng Bo
- 云斑天牛的交配产卵行为
- Mating and Oviposition Behavior of Batocera horsfieldi
- 林业科学, 2011, 47(6): 88-92.
- Scientia Silvae Sinicae, 2011, 47(6): 88-92.
-
文章历史
- 收稿日期:2010-06-22
- 修回日期:2010-10-25
-
作者相关文章



2. 贵州大学昆虫研究所 贵州山地农业病虫害重点实验室 贵阳 550025;
3. 四川省德阳市林业局 德阳 618000
2. Guizhou Key Laboratory for Plant Pest Management of Mountain Region Institute of Entomology, Guizhou University Guiyang 550025;
3. Forestry Bureau of Deyang of Sichuan Province Deyang 618000
云斑天牛(Batocera horsfieldi)是短周期用材林和经济林的重要钻蛀性害虫。杨树(Populus spp.)因云斑天牛蛀害,轻则树势衰弱,重则全株枯死; 同时还造成材质损害,给生产造成极大损失(梅爱华等,1998)。近年来,在我国北方核桃(Juglans regia)主产区的核桃树也严重受害,虫株率70%以上,成为发展核桃的重要制约因素(王绍林等,2004)。李建庆等(2009)分析确定云斑天牛在中国有害生物风险性分析PRA值为2. 04,属高度危险性林业有害生物。成虫是云斑天牛唯一有行为导向的虫态,其行为导向是对嗜食寄主呈明显的趋性,这一趋性的源动力在于满足生殖活动对营养的需要(严敖金等,1997)。梁潇予等(2008)发现云斑天牛成虫对法国冬青(Viburnum awabuki)和光皮桦(Betula luminifera)有较强的嗜食性。李娟等(2008)在田间调查中发现云斑天牛成虫多集中在蔷薇科植物上补充营养。而补充营养对云斑天牛的繁殖起着决定性的作用,取食不同的寄主对成虫的生殖力有显著影响,以野蔷薇(Rosa multiflora)饲养云斑天牛的产卵量为每头13. 8粒,而以旱柳(Salix matsudana)、杨树及枫杨(Pterocarya stenoptera)饲养的天牛则不产卵(高瑞桐等,1998)。
天牛交配前一般没有复杂的求爱行为,雌雄个体聚集后易发生交配,但每次交配持续时间差异很大(Hanks,1999; 贺萍等,1993)。在对天牛的行为学研究中,贺萍等(1993)发现光肩星天牛成虫的交配行为分为徘徊、接近、就位、舔翅和交配5个阶段。杨洪等(2007)认为松褐天牛(Monochamus alternatus)一次完整的交配过程包括相遇抱对、插入输精和配后保护3个阶段。尹新明(1996)对狭胸天牛(Philus antennatus)的成虫相遇至交配的各个阶段进行详细的观察,发现雄虫活跃,在短距离内能识别雌虫,交配过程为追逐、上背、定位、曲腹、插入输精。本文通过对云斑天牛成虫交配及产卵行为的研究,旨在掌握其行为活动规律、丰富昆虫生殖行为学知识,为云斑天牛的行为控制技术提供参考。
1 材料与方法 1.1 供试虫源2010年4月下旬,于四川省德阳市罗江县毛白 杨(Populus tomentosa)上采集刚羽化未交配的云斑天牛成虫,鉴别雌雄虫后分别放入60 cm × 60 cm × 60 cm的不锈钢网养虫笼内,用水培光皮桦枝条室温下饲养备用。已交配雄虫腹部绒毛磨损,形成明显的交配斑; 已交配雌虫为背部绒毛磨损,形成明显交配斑(嵇保中等,1996)。
1.2 室内交配试验用白色记号笔分别在雌虫腹部和雄虫背部标注不同数字,以此标记同期羽化的云斑天牛成虫4对(雌雄大小一致,无畸形,羽化出孔后饲养8天),置于60 cm × 60 cm × 60 cm的不锈钢网养虫笼内,以水培光皮桦1 ~ 2年生枝条饲养,并放入长1 m,直径为15 cm的新鲜杨树木段作为产卵诱木。2天更换1次光皮桦枝条和产卵诱木。观察记录云斑天牛配偶抱对、输精及配后保护的行为与时间。每天8:00—20:00进行观察,20:00后取出笼内雄虫单独饲养。连续观察4天,试验重复4次。
1.3 野外交配试验2010年4月下旬在四川省德阳市罗江县杨树林内,每天8:00—20:00对在补充营养寄主野蔷薇上交配的云斑天牛进行观察,记录云斑天牛配偶抱对、输精及配后保护的行为与时间,连续观察10天。
1.4 成虫取食不同植物对产卵量的影响采用旱柳、核桃、法国冬青、光皮桦、杨树1 ~ 2年生小枝,将5种枝条分别放入60 cm × 60 cm × 60 cm的不锈钢养虫笼内,插入盛有清水的三角瓶中,以杨树木段为产卵诱木。每2天更换1次枝条和清水。养虫笼内放入同期羽化并饥饿24 h的云斑天牛成虫8头(雌雄比为1: 1),每天8:00—20:00进行观察,连续观察10天。计算取食枝皮面积和产卵数量,试验重复4次。
1.5 成虫对产卵诱木选择试验将直径5 ~ 10 cm、长30 cm的产卵诱木(核桃、杨树、法国冬青、光皮桦、野蔷薇),与同期羽化后饲养8天的云斑天牛雌雄成虫4对,放入60 cm × 60 cm × 60 cm的不锈钢养虫笼内,以水培光皮桦1 ~ 2年生枝条饲养。连续观察10天,记录产卵诱木上的产卵数量。试验重复4次。
1.6 成虫产卵进度试验在60 cm × 60 cm × 60 cm的不锈钢养虫笼内放入羽化出孔后饲养8天的20头雌虫和20头雄虫,以水培光皮桦1 ~ 2年生枝条饲养。每2天更换一次新鲜光皮桦枝条和杨树产卵木,记录其上的产卵数,连续观察30天。
1.7 数据分析试验所得数据用SPSS 13. 0进行统计分析,采用Microsoft Office Excel 2003绘图,成虫交配行为室内观察与野外观察结果采用两样本配对t检验比较,取食面积和产卵量采用Duncan's多重比较。
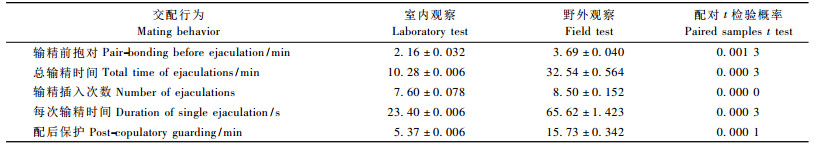
2 结果与分析 2.1 云斑天牛成虫交配行为通过室内和野外观察发现,云斑天牛一次完整的交配过程分为相遇抱对、插入输精和配后保护3个阶段,其中会有多次插入输精。雄虫在交配中占主导地位,在爬行中遇到雌虫后用触角扫探雌虫进行识别,并立刻进行抱对,没有明显的求偶阶段。室内试验共观察到云斑天牛交配169次,其中成功完成交配的121次,有配后保护的也是121次。一次完整的交配过程平均需要17. 81 min,其中输精前抱对平均时间为2. 16 min,总输精时间平均为10. 28 min,交配过程中每次输精插入时间平均为23. 4 s,配后保护时间为5. 37 min (表 1)。在连续10天的野外观察中,共观察到有完整交配过程的云斑天牛5对,野外交配行为主要发生在傍晚。所观察到的野外交配行为都发生在补充营养寄主上。其中输精前抱对平均时间为3. 69 min,总输精时间平均为32. 54 min,交配过程中每次输精插入时间平均为65. 62 s,配后保护时间为15. 73 min。
|
|
通过对16对云斑天牛4天内交配情况的观察发现,云斑天牛4天内每头雄虫平均交配次数为12. 94次,最少只有1次,最多的达33次,说明不同雄性个体所获得的交配机会差异很大(图 1)。如果继续延长观察时间,雄虫平均交配次数可能还会增多。雌虫平均交配次数为12. 94次,最少交配6次,最多28次。雌虫交配的个体间也存在较大差异(图 2)。

|
图 1 云斑天牛雄虫交配次数分布 Figure 1 Copulatory frequency distribution of B. horsfieldi males |

|
图 2 云斑天牛雌虫交配次数分布 Figure 2 Copulatory frequency distribution of B. horsfieldi females |
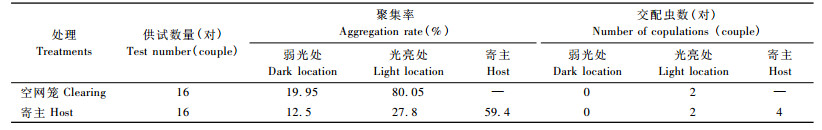
云斑天牛在不同环境中的聚集、交配情况见表 2。放入网笼内的雌、雄成虫先各自不定向地爬行或做短距离的飞行,而后会向上攀爬或向光亮处运动。当设置有寄主植物时,雌、雄成虫会趋向在寄主植物上聚集。寻偶、交配主要发生在寄主植物上。
|
|
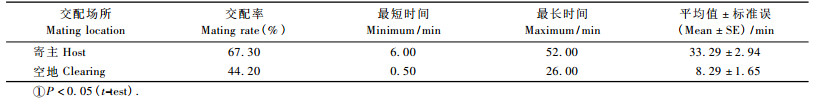
不同交配场所16对云斑天牛交配率、平均交配持续时间如表 3。云斑天牛在寄主植物和空地上的交配率分别为67. 30%和44. 20%;而在寄主植物上的平均交配时间为33. 29 min显著高于在空地上的平均交配时间8. 29 min (P<0. 05)。
|
|
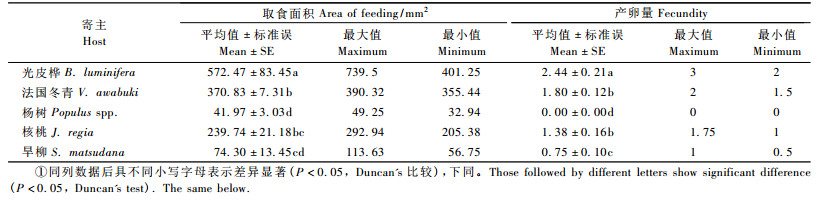
对取食5种不同植物的枝皮面积和产卵数量的观察结果见表 4,云斑天牛取食光皮桦的枝皮面积最大,平均每天每头达到572. 47 mm2,其次是法国冬青的370.83 mm2,杨树的最小为41.97 mm2。取食光皮桦和法国冬青的云斑天牛平均每天每头产卵数量最大,分别为2. 44和1. 8粒,取食杨树的不产卵。多重比较结果表明,云斑天牛对光皮桦的取食量和取食后的产卵量同其余4种树种相比均达到显著性差异(P<0. 05)。
|
|
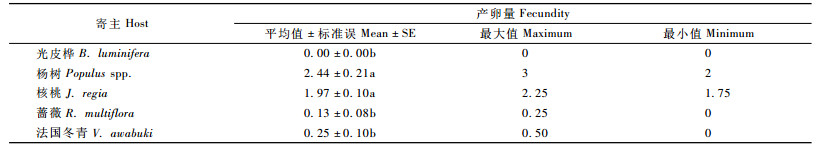
在对5种产卵诱木的选择上,云斑天牛在杨树上的产卵数最多,平均每天每头达到2.44粒,其次为核桃1.97粒,在光皮桦上没有产卵情况,在杨树和核桃上产卵数量明显多于蔷薇、法国冬青和光皮桦,达到显著性差异(P<0. 05) (表 5)。
|
|
通过30天的产卵进程观察,云斑天牛共产卵510粒。第10天到14天之间雌虫产卵出现高峰期,达126粒,占全部产卵量的24. 7%,第26天后没有产卵情况发生(图 3)。

|
图 3 云斑天牛室内产卵情况 Figure 3 Oviposition of B. horsfieldi in the laboratory |
在天牛交配行为的研究中,许多学者把插入内囊输精到输精完成作为一次完整的交配过程(Hanks,1999; 张永慧等,2006)。实际上天牛的交配是一个复杂的过程,它涉及输精过程前后的多个阶段(杨洪等,2007)。本研究发现云斑天牛交配的完整过程分为输精前抱对、插入输精和配后保护3个阶段。云斑天牛没有明显的求偶阶段,雄虫发现雌虫后会用下颚须和下唇须不断地舐触雌虫的前胸背板和鞘翅,抱握骑乘,弯曲腹部,如果雌虫不反抗,则顺利达成交配。
云斑天牛野外和室内交配过程一致,但在野外交配过程中,各阶段的平均耗时都显著长于室内。在野外没有人为干扰的情况下,影响云斑天牛交配的主要因素是温度,低温时期交配主要集中在10:00—14:00时,高温时则主要在傍晚和黎明(严敖金等,1997)。室内交配试验中,云斑天牛雄虫因有足够多的雌虫进行交配,雄虫会对交配对象作出选择,这可能也是室内交配各阶段用时低于野外观察的原因之一。
云斑天牛羽化出孔后成虫寿命较长,雄虫28 ~ 82天,众数历期40 ~ 45天; 雌虫30 ~ 146天,众数历期60 ~ 65天(严敖金等,1997)。在此期间雌雄成虫均需要补充营养。云斑天牛正是依靠寄主植物的诱集作用,完成长距离的聚集。而在寄主树木上的栖息活动中,雌、雄成虫对寄主树木特定区域微生境的共同趋性使得两性接近,从而完成近距离的聚集; 云斑天牛成虫的交配行为大都是在雄虫接触到雌虫后,便立即快速爬向雌虫。在交配活动中,雌虫存在接触性信息素对雄虫的接触性刺激作用非常重要(杨远亮,2008),雄虫因直接接触雌虫而引发性兴奋,追逐和骑乘抱握雌虫、曲腹,并最终插入内囊完成交配。研究中发现寄主植物不仅在云斑天牛两性间远距离通讯中起着重要的作用,并且其存在与否对云斑天牛的交配成功率有着显著的影响。研究表明,寄主植物气味能增强昆虫对性信息素的反应(杜永均等,1994; Hansson et al., 1989; Landolt et al., 1994; 杨桦等,2010)。云斑天牛是否也存在这一现象,还有待进一步研究。
云斑天牛成虫对光皮桦和法国冬青有较强的嗜食性,并且取食后的产卵数量最大。高瑞桐等(1995)研究证明植物体内糖含量与云斑天牛成虫的取食量、雌虫寿命、产卵量成正相关。这揭示光皮桦和法国冬青体内的糖含量能满足云斑天牛的自身需求。而云斑天牛取食杨树后不产卵这个结果同高瑞桐等(1995)、孙巧云等(1991)的研究一致。云斑天牛对产卵诱木的选择主要以杨树和核桃为主,法国冬青和野蔷薇有少量刻槽情况,成虫对产卵寄主的选择主要是寄主气味对嗅觉上的吸引,还是其他信息素的吸引及视觉因素或者是这些因素的共同作用还有待进一步系统的研究。
云斑天牛室内产卵呈周期性波动,每隔5 ~ 6天出现一个产卵周期,这同夏剑萍等(2005)对云斑天牛分批产卵,成虫交尾1次产卵1次,雌虫一生要产5 ~ 6次卵,每次产卵的周期为8 ~ 10天的结论基本一致。通过对云斑天牛产卵观察发现,成虫活动场所大致呈现在成、幼期寄主间转换的昼夜节律,白天主要进行取食和交配,产卵则在夜间进行(严敖金等,1997)。
杜永均, 严福顺, 韩心丽, 等. 1994. 大豆蚜嗅觉在选择寄主植物中的作用[J]. 昆虫学报, 37(4): 385-392. |
高瑞桐, 王宏乾, 徐邦新, 等. 1995. 云斑天牛补充营养习性及与寄主树关系的研究[J]. 林业科学研究, 8(6): 619-623. |
高瑞桐, 郑世锴. 1998. 利用成虫取食习性防治3种杨树天牛[J]. 北京林业大学学报, 20(1): 43-48. |
贺萍, 黄竞芳. 1993. 光肩星天牛成虫的行为[J]. 昆虫学报, 36(1): 51-55. |
嵇保中, 钱范俊, 严敖金. 1996. 云斑天牛研究方法的改进[J]. 森林病虫通讯, (1): 45-46. |
李建庆, 杨忠岐, 梅增霞, 等. 2009. 云斑天牛的风险分析及其防治控制对策[J]. 林业科学研究, 22(1): 148-153. |
李娟, 王满囷, 张志春, 等. 2008. 云斑天牛成虫对植物气味的行为反应[J]. 林业科学, 44(6): 168-170. DOI:10.11707/j.1001-7488.20080628 |
梁潇予, 杨伟, 杨远亮, 等. 2008. 云斑天牛对补充营养寄主的选择性[J]. 昆虫知识, 45(1): 78-82. DOI:10.7679/j.issn.2095-1353.2008.016 |
梅爱华, 陈京元, 吴高云, 等. 1998. 江汉平原杨树害虫种类调查、发生原因及主要害虫防治对策[J]. 森林病虫通讯, (2): 35-39. |
孙巧云, 赵自成. 1991. 云斑天牛初步研究[J]. 江苏林业科技, (2): 22-25. |
王绍林, 王宏琦, 夏明辉, 等. 2004. 核桃树云斑天牛的发生规律与防治技术[J]. 中国果树, 3(2): 11-13. |
夏剑萍, 戴均华, 刘立德, 等. 2005. 云斑天牛研究进展[J]. 湖北林业科技, 132(2): 42-44. |
严敖金, 嵇保中, 钱范俊, 等. 1997. 云斑天牛Batocera horsfieldi(Hope)的研究[J]. 南京林业大学学报, 21(1): 1-5. |
杨洪, 王进军, 赵志模, 等. 2007. 松褐天牛的交配行为[J]. 昆虫学报, 50(8): 807-812. |
杨桦, 杨茂发, 杨伟, 等. 2010. 竹横锥大象对寄主及虫体挥发物的行为和触角电位反应[J]. 昆虫学报, 53(3): 286-292. |
杨远亮. 2008. 云斑天牛成虫聚集、交配行为的初步研究. 雅安: 四川农业大学硕士学位论文.
|
尹新明. 1996. 狭胸天牛生殖行为的研究[J]. 河南农业大学学报, 30(4): 347-349. |
张永慧, 郝德君, 王焱, 等. 2006. 松墨天牛成虫交配与产卵行为的观察[J]. 昆虫知识, 43(1): 47-49. DOI:10.7679/j.issn.2095-1353.2006.010 |
Hanks L M. 1999. Influence of the larval host plant on reproductive strategies of cermbycid beetles[J]. Ann Rev Entomol, 44: 483-505. DOI:10.1146/annurev.ento.44.1.483 |
Hansson B S, Van Der Pers J N C, Lofquist J. 1989. Comparison of male and female olfactory cell response to pheromone compound and plant volatiles in turaip moth, Agrotis segetum[J]. Physiol Entomol, 14: 147-155. DOI:10.1111/j.1365-3032.1989.tb00946.x |
Landolt P J, Heath R R, Miller J G, et al. 1994. Effects of host plant Gossypium hirsutum L.on sexual attraction of cabbage looper moths Trichoplusia ni[J]. J Chem Ecol, 20(11): 2959-2975. DOI:10.1007/BF02098402 |