 2011, Vol. 47
2011, Vol. 47文章信息
- 卢军, 李凤日, 张会儒, 张守攻
- Lu Jun, Li Fengri, Zhang Huiru, Zhang Shougong
- 帽儿山天然次生林主要树种冠长率模型
- A Crown Ratio Model for Dominant Species in Secondary Forests in Mao'er Mountain
- 林业科学, 2011, 47(6): 70-76.
- Scientia Silvae Sinicae, 2011, 47(6): 70-76.
-
文章历史
- 收稿日期:2009-07-29
- 修回日期:2010-10-25
-
作者相关文章


2. 东北林业大学林学院 哈尔滨 150040;
3. 中国林业科学研究院林业研究所 北京 100091
2. Forestry College, Northeast Forestry University Harbin 150040;
3. Research Institute of Forestry, CAF Beijing 100091
冠长率(crown ratio)是指树冠的长度与树木总高度的比率,是一种无量纲的测度。树冠的大小是树冠活力一个很重要的测度(Assmann,1970; Spurr et al., 1980; Valentine et al., 1994)。树冠的测量对评价树干的木材质量和商业价值起到重要的作用(Kershaw et al., 1990)。Monserud(1975)认为冠长率是一个树木利用可获得的资源来促进生长的能力指标,Daniels等(1979)认为树冠长度和冠长率都可以反映疏开木利用可获得资源的潜能。
单木生长模型一般被划分为利用树冠因子来计算竞争(用树冠的冠幅、面积或者体积)和利用树冠的生长空间(用冠长率、树冠长度)来计算树木的生长潜力(Monserud,1975)。几十年来,冠长率被成功地用于预估长时期的树木生长量(Monserud,1975; Daniels et al., 1975; Monserud et al., 1996)。Monserud等(1996)发现用对数形式来预估冠长率,系数高度显著,并且对奥地利森林中所有树种断面积生长量的预测是正向的。Monserud(1975)使用一种冠长率异速生长模型的变形来修正美国Wisconsin北方硬阔林潜在的树高和直径生长,树高异速生长方程的决定系数在12个树种中有11个显著,在直径生长方程中,有9个显著。Wykoff(1990)发现在北部落基山脉11个混交针叶树种的断面积生长量预测中,冠长率是一个高度显著的预测因子。Daniels等(1975)在美国东南部使用冠长率来预测火炬松(Pinus taeda)人工林树高生长量和用树冠长度来预测直径生长量。Shifley(1987)发现在美国中部22个树种组的断面积生长量预测中,有19个树种组的冠长率是显著的预测因子。在所有这些研究中,每一个显著的冠长率的系数都是正向的,冠长率的增加会导致断面积生长的增加。
国内对于冠长率模型,只有少量的学者在研究中利用到了冠长率这一因子,如杜纪山(1991)研究了油松(Pinus tabulaeformis)的冠长率与干形之间的关系,指出冠长率和平均高对油松的干形影响不显著。而真正建立冠长率相关的模型则极少。
由于树木的树冠反映了过去时间的累积的竞争水平,因此本研究的目标是使用森林调查数据,依据冠长率与树冠大小和林分因子之间的异速生长关系,建立单木冠长率模型,同时分析不同林分因子对冠长率的影响,可为建立大地域林分生长模型或者单木生长模型获得基础的预测指标。在森林经营中,尤其是在我国实施森林分类经营后,研究冠长率、分析冠长率对树木生长的影响,为用材林的经营提供可靠的理论依据是很有意义的。
1 研究地概况本研究选择具有典型黑龙江省森林林分特点的东北林业大学帽儿山实验林场为研究地,该地区具有典型的次生林结构。帽儿山实验林场位于黑龙江省东南部,45°20'—45°25' N。总面积26 496 hm2,南北长30 km,东西宽20 km,共151个林班。该地区属温带季风气候,但是具明显的大陆性,四季分明,冬长夏短。地带性土壤为暗棕壤,有机质含量、各种化学元素、微量元素含量都较高。植被属于长白山植物区系,是由地带性顶级植被阔叶红松(Pinus koraiensis)林经人为干扰破坏后形成的较典型的东北东部天然次生林。
2 数据收集与整理2007年共设置了30块天然次生林标准地,位于帽儿山实验林场的中林和跃进施业区,所选的标准地所在林分均是未经过间伐、生长正常、具有代表性的次生林,每块标准地都有GPS定位,在2个施业区的位置分布如图 1所示。

|
图 1 标准地位置示意 Figure 1 Location of sample plots |
标准地规格:根据地形和林相特点,设置不同面积的标准地,平均为20 m × 50 m,最大为30 m ×70 m,标准地状况见表 1。
|
|
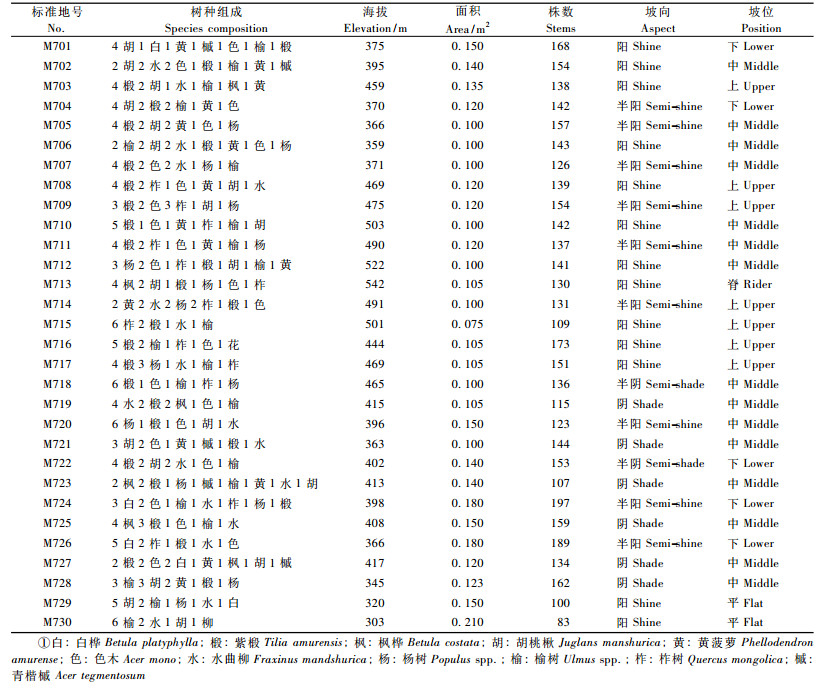
把这30块不同年龄、不同立地条件、不同密度的天然次生林标准地分别编号为M701,M702,… M730。对每块固定标准地完成上述因子测定并建立数据库,各标准地基本概况见表 2。在所设置的30块天然次生林标准地中,共测得样木4 237株,剔除枯死木和亚乔木,有效的数据为3 628株,其基本情况见表 2。
|
|
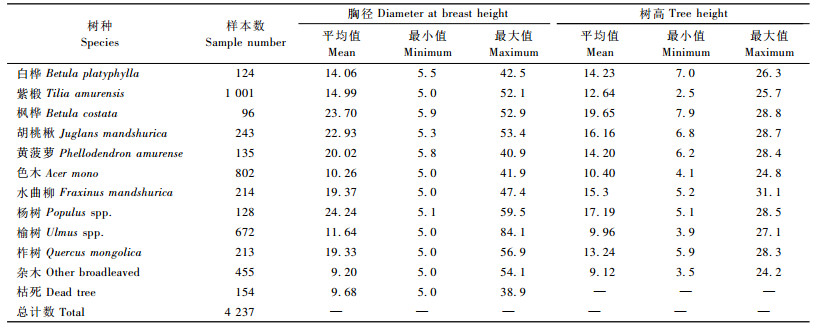
为了建立次生林阔叶树种冠长率的模型,需要把这些数据分成计算数据和检验数据,比例分别约为80%和20%。
3 模型设计与参数估计 3.1 模型设计由冠长率(CR)的定义,它是在0(没有树冠)到1(全树冠)范围之内的。因此排除了模型中超过范围的和由极限数据而得出的不合逻辑的结果。
冠长率是受林分密度和林分内树木的种群位置影响的。散点图和数据分析结果表明:冠长率和林分密度之间的关系表现出了3种等级。树木在低密度的林分中一般有一个大的冠长率,但是在这个范围内,密度和冠长率之间没有联系(Hasenauer et al., 1996)。随着密度的增加,树冠衰退加强。当活树冠减小至CR = 0. 3以下时,冠长率和林分密度之间再次表现出随机变化。虽然这种关系因树种而异,但是会出现相似的趋势。这种表现是一个饱和曲线的特征,尤其是Logistic方程能很好地模拟这个特征:
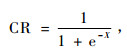
|
(1) |
式中:CR是冠长率; X是输入变量和估计参数的线性组合。
Logistic方程是在限制生长的差分方程中产生的,一开始由Verhulst引进到人口增长,后来被Robtson和Ostwald改进到器官的生长预测中(Finney,1971)。Neter等(1970)把Logistic模型引入到单木枯损模型的预估,而Hatch (1980)和Hasenauer等(1996)使用它来预测冠长率。
在本研究中,X可以描述为林木大小变量、竞争变量和立地变量的函数,因此X可以表示为:
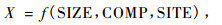
|
(2) |
式中:SIZE表示林木大小变量; COMP表示竞争变量; SITE表示立地变量。
3.1.1 林木大小变量选择冠长率的相关因子时,树木大小变量、树高(H)和胸径(D)是基本的形式,一株树木的树高/胸径比(H /D)是树干很重要的一个因子,是和冠长率相关的,树干和冠长率都受林分密度的影响。树干部分较长的树木会比树干矮小的树木有更高的冠长率(Abetz,1976)。树木的大小变量可以由下式表示:
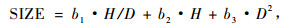
|
(3) |
式中: H /D是树高胸径之比。
3.1.2 竞争变量在森林调查中往往很少测量树木的空间坐标信息,因此竞争变量不包括单木的空间信息,也就是与距离无关。另外,冠长率模型应该同时适用于同龄林和异龄林的林分条件,因此,密度测量需要一个与年龄无关的收获表(如立木度水平)来确定。密度是影响体现于基于直径的大树的断面积(BAL),它表明了林分内一株树木的群落等级和相对的优势程度(Wykoff,1990),Krajicek等(1961)提出的树冠竞争因子(CCF)。CCF的优点是它一般与林分的立地条件和年龄无关。BAL的优点是它可以很好地表现各种类型的间伐措施下的竞争的影响(Wykoff,1990)。因此,竞争变量可以表达为:
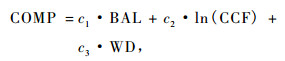
|
(4) |
式中:BAL是比目标树的直径大的树木的每公顷断面积; CCF是树冠竞争因子; WD为冠幅。
CCF是用疏开木冠幅方程(Hasenauer et al., 1994)计算的,它只用直径大于10 cm的树木来计算,因此在复层林分的更新层中可以剔除小树。笔者用CCF的对数来减少大的CCF估计值的抽样变化影响。
3.1.3 立地变量立地因子是与标准地相关的变量,可以反映地形因子,在同一标准地中,所有样木的立地都是相同的。立地变量可以表示为:

|
(5) |
式中:ELEV是百米形式的海拔; SL是坡度的正切值; AZ是坡位的弧度值。
海拔的平方和坡度的变形是为了考虑优化和最大化海拔和坡度的影响,这种坡度-坡位变形是Stage(1976)提出的。全部的模型形式如下:
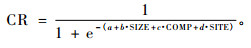
|
(6) |
本研究用Statistica 7. 0中的非线性回归来估计Logistic方程的参数,显著水平α = 0. 05。
对公式(6)两边去对数后,用线性回线性化模型来进行估计:
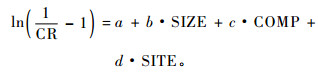
|
(7) |
这种线性化对于冠长率的极值0和1是错误的。冠长率为0的树木是死亡的,冠长率为1表示全树冠,这在现实林分中很罕见。在实际的拟合过程中,这样的数据都被剔除。
用每个变量组(SIZE,COMP和SITE)对变动解释的百分比来评价它们对模型的贡献程度,用以下的方法来计算每个变量的多重决定系数(Ri2)。每一个变量解释的变异程度可以表示为估计的回归系数的乘积,也就是因变量和自变量的偏差的交叉乘积之和。统计变量的残差平方和,并计算解释变动和总变动之间的比率,模型中每个变量的非调整的多重决定系数(Ri2)可以表示为:
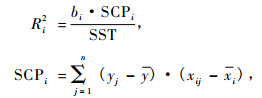
|
(8) |
式中: yj是因变量y的第j个观测值(1到n),y是因变量的平均值; xi是自变量的第i个值,xij是第i个自变量的第j个观测值; xi为第i个自变量平均值; Ri2为自变量i的多重决定系数; 为变量i的回归估计系数; SCPi是偏差的交叉乘积之和; SST是yi和y之间的离差平方和。
3.3 模型的检验本研究利用检验样本数据,通过以下几种偏差统计量作为比较和评价模型预测能力的指标:
1) 平均偏差(mean error)
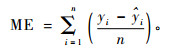
|
(9) |
2) 均绝对偏差(absolute mean error)
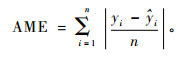
|
(10) |
3) 均相对偏差(relative absolute error)
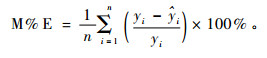
|
(11) |
4) 均相对偏差绝对值(relative absolute mean error)
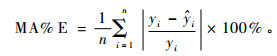
|
(12) |
5) 模型的预估精度(P%)
利用独立检验样本数据,通过实测值yi和模型预估值xi之间建立一元线性回归方程:
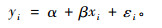
|
由检验数据(xi,yi) (i = 1,2… n),采用最小二乘法估计回归系数α和β的估计值a和b,并计算出回归标准差(Sy.x),回归标准误(Sy),误差限(△)及相对误差限(E%) :
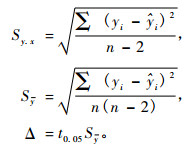
|
模型的预估精度(P%) :

|
(13) |
式中: yi为实测值; 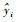
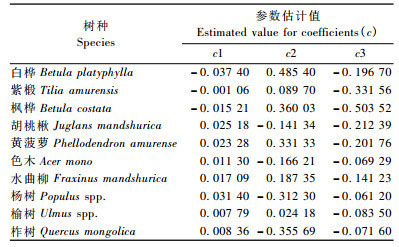
经过拟合公式(7),得出每个树种的参数估计结果见表 3~5。
|
|

|
|
|
|

公式(7)中所有的拟合结果表明:对于每个树种的变量解释百分比,从最小的色木的12. 881 3%到白桦的42. 116 8%,对于帽儿山次生林区分布最多的树种紫椴的解释变量百分比是17. 403 2% (表 6)。天然次生林结构复杂,层次多变,类型多样,因此本研究所建立的帽儿山林区10个主要树种的冠长率模型的决定系数都不算很高。
|
|
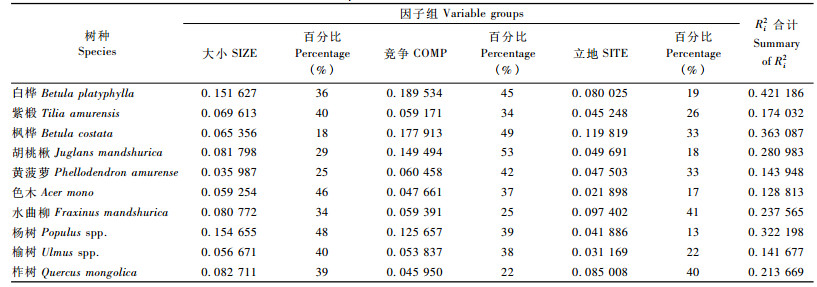
林木大小、竞争和立地这3个因子组对于每个树种所起到的作用不一样。从表 6可知:除了枫桦、黄菠萝、水曲柳和柞树外,立地因子组对各树种的冠长率模型影响较小,最小的是杨树13%,说明坡度、坡向和坡位等立地因子的差异性对这6个树种的冠长率影响不大。影响较大的这4个树种也和本身的生物学特性相关,它们的生长、分布都受立地条件的影响。林木之间的竞争对白桦、枫桦、胡桃楸的影响显著,竞争因子组所占的决定系数百分比超过了40%,也和这4个树种的特点密不可分,它们的树冠都相对较大,与相邻木之间的光线竞争就比较强烈。林木的大小因子组中,紫椴、色木和杨树3个树种对大小因子的反应比较显著,所占决定系数百分比都超过了40%,说明树木的胸径、树高对它们的冠长率的影响较大,尤其是色木处于上层的林木的胸径必然就会较大,也是获得了较强的竞争能力。总的来说,3个因子组中林木的大小和竞争对树木的冠长率影响较大,而立地条件影响相对较小。
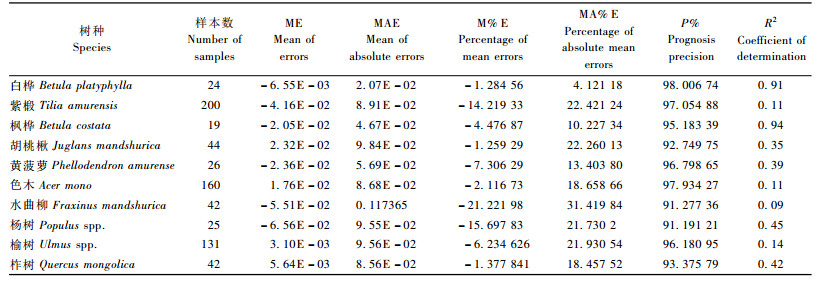
4.2 冠长率模型的检验模型的独立性检验主要是计算几种误差值及其百分比,对公式(7)所建立的10个树种的冠长率模型进行检验,模型的检验结果见表 7。
|
|
用来自30块标准地的10个树种的检验数据来检验模型的优良程度,这些数据在帽儿山地区具有代表性,代表了调查区域各种林分条件下树木的生长情况。本研究的测量过程比较困难的地方是阔叶树种第1活枝高度的确定,也就是到树冠基部的高度,这在针叶树种中是比较容易的,因为多数针叶树种具有轮枝生长现象,因此,活枝高度很容易观测。但是阔叶树种的情况大不相同,在树干的下部也会生长一些小的侧枝,并且没有规律,或者有的活枝条间断的分布在树干上,这对确定第1活枝高度带来了很大的困难。本研究使用超声波测高器来确定树高和第1活枝的高度,每个测量者对第1活枝的判断是不同的,但是遵循的原则就是尽可能把树干上第1个活的较大的枝作为第1活枝,从而测得地面到树冠基部的高度,其他的树干上往下的小侧枝生活力低,可能会在以后的生长过程中死亡,因此不能作为第1活枝。在测量的过程中,对第1活枝高度的测定的不确定性导致了冠长的计算误差,因此,所拟合的冠长率模型会有过估或低估的现象发生。
表 7表明各个树种的冠长率模型的检验结果。所有的10个树种的预估精度都超过了91%,白桦最高达到了98%以上。几种偏差值都较小,尤其是ME和MAE值很小。对于所构造的直线回归的决定系数R2,多数树种都较低,只有白桦和枫桦较高,超过了0. 9。胡桃楸、黄菠萝、杨树和柞树的R2值在0. 35 ~ 0. 45之间,效果比较好。而紫椴、色木、榆树和水曲柳的R2值都较低,在现实林分中,这4个树种的冠长变动范围非常大,尤其是紫椴和色木,处于林分下层时,冠长非常大,但在林分上层时,冠长较小,这为模拟工作带来难度。
5 结论与讨论采用3个变量组(SIZE,COMP,SITE)建立单木生长模型是一种把树木的林学意义和数学的模拟结合到一起的有效途径,本研究基于此建立的单木冠长率模型能够合理地模拟黑龙江省帽儿山林场次生林内10个阔叶树种的冠长率的规律。10个树种的冠长率模型并不是一致的,这也是和次生林的立地条件有关系,复杂、多样化的立地和生境使阔叶树种的生长也变得难以预测,因此对不同冠长率的树种应采取不同的间伐措施。类似的研究也表明,冠长率在不同的树种中有不一致的表现,比如在澳大利亚云杉(Picea asperata)、冷杉(Abies fabri)和欧洲赤松(Pinus sylvestris)随冠长率改变而显著影响生长的预测,橡木(Quarcus)、山毛榉(Fagus sylvatica)和黑松(Pinus thunbergii)则是最不显著的,Abetz (1974)在未来间伐方针里建议,对橡木应有最宽的间伐距离,山毛榉较小,欧洲云杉(Picea abies)、欧洲赤松和冷杉最小。
Monserud(1975)认为对于欧洲云杉、冷杉和黑松,林木大小这个变量组与竞争变量组的重要性是相当的; 竞争变量组对欧洲赤松和黑松是比林木大小变量组要重要的。而在本研究中,林木大小、竞争和立地这3个因子组对于每个树种所起到的作用是不一样的。不同的树种的生物学特性决定了变量组的影响作用,比如枫桦、黄菠萝、水曲柳和柞树对立地因子的反应。而大小因子则表现为树冠、胸径的综合作用,进一步反映出光合作用对树木竞争能力的影响。总的来说,3个因子组中林木的大小和竞争对树木的冠长率影响较大,而立地条件相对较小。坡度、坡向、坡位和海拔等立地因子是大环境的变量,需要大量的样本数据才能得出规律,因此,本研究限于数据量较少,而不能分析冠长率与这些因子的关系。
杜纪山. 1991. 油松相对干形与树冠率及平均高的关系[J]. 陕西林业科技, (2): 53-58. |
Abetz P. 1976. Beitrage zum Baumwachstum Der h/ d Wertmehr als ein Schlankheitsgrad[J]. Forstwirtsch Holzwirtsch, 31(19): 389-393. |
Assmann E.1970.The The Principles of Forest Yield Studies.Pergamon Press, Oxford, 506.
|
Daniels R F, Burkhart H E, Spittle G D, et al.1979.Methods for modeling individual tree growth and stand development in seeded loblolly pine stands.Publ.FWS ~ 1 ~ 79, College of Forestry and Wildlife Resources, Virginia Technical Institute, Blacksburg, 50.
|
Daniels R F, Burkhart H E.1975.Simulation of individual tree growth and stand development in managed loblolly pine plantations on cutover, site-prepared areas.Pub1.FWS5-75, College of Forestry and Wildlife Resources, Virginia Technical Institute, Blacksburg, 69.
|
Finney D J. 1971. Probit Analysis[M]. 3rd ed. London: Cambridge University Press: 333.
|
Hasenauer H, Monserud R A. 1996. A crown ratio model for Austrian forests[J]. Forest Ecology and Management, 84(1/3): 49-60. |
Hatch C R. 1980. Modeling tree crown size using inventory data[J]. Mitt Forstl Bundes-Versuchsanst Wien, 130: 93-97. |
Kershaw J A, Jr Maguire D A, Hann D W. 1990. Longevity and duration of radial growth in Douglas-fir branches[J]. Can J For Res, 20(11): 1690-1695. DOI:10.1139/x90-225 |
Krajicek J E, Brinkman K A, Gingrich S F. 1961. Crown competition: a measure of density[J]. For Sci, 7(1): 35-42. |
Monserud R A, Sterba H. 1996. A basal area increment model for individual trees growing in even and uneven-aged forest stands in Austria[J]. For Ecol Manage, 80(1/3): 57-80. |
Monserud R A.1975.Methodology for simulating Wisconsin Northern hardwood stand dynamics.Ph.D.Thesis, University of Wisconsin, Madison, 156.
|
Neter H, Maynes S E. 1970. On the appropriateness of the correlation coefficient with a 0, 1 dependent variable[J]. J Am Stat Assoc, 65(330): 501-509. DOI:10.1080/01621459.1970.10481099 |
Shifley S R.1987.A generalized system of models forecasting central states tree growth.Res Pap NC ~ 279, USDA Forest Service, 10.
|
Spurr S H, Barnes B V. 1980. Forest Ecology[M]. 3rd ed. New York: John Wiley: 527.
|
Stage A R. 1976. An expression for the effect of slope, aspect and habitat type on tree growth[J]. For Sci, 22(4): 457-460. |
Valentine H T, Baldwin V C, Gregoire T G, et al. 1994. Surrogate for foliar dry matter in 1oblolly pine[J]. For Sci, 40(3): 576-585. |
Wykoff W R. 1990. A basal area increment model for individual conifers in the Northern Rocky Mountains[J]. For Sci, 36(4): 1077-1104. |