 2011, Vol. 47
2011, Vol. 47文章信息
- 谭小梅, 金国庆, 邵纪清, 余家中, 陆名革, 周志春
- Tan Xiaomei, Jin Guoqing, Shao Jiqing, Yu Jiazhong, Lu Mingge, Zhou Zhichun
- 马尾松巢式交配子代生长和木材基本密度遗传分析
- Genetic Analysis of Growth and Wood Density of Progeny of Nested Mating Pinus massoniana
- 林业科学, 2011, 47(6): 30-35.
- Scientia Silvae Sinicae, 2011, 47(6): 30-35.
-
文章历史
- 收稿日期:2010-04-14
- 修回日期:2010-06-21
-
作者相关文章



2. 重庆市林业科学研究院 重庆 400036;
3. 浙江省淳安县富溪林场 淳安 311700
2. Chongqing Academy of Forestry Chongqing 400036;
3. Fuxi Forest Farm of Chun'an County, Zhejiang Province Chun'an 311700
杂交育种是植物遗传改良的一种有效手段和主要途径, 是目前最有成效的育种方法之一, 在植物遗传改良实践中已得到了广泛应用(马常耕, 1995; King et al., 1997), 其中亲本的选配是杂交育种成败的先决条件(张爱民, 1994)。植物育种实践证明, 杂交后代的表现与其亲本本身的表现往往不一致, 且不同亲本间的组配能力也不相同, 因此了解亲本在杂交组合中的实际表现, 对于指导杂交育种工作具有非常重要的意义(孔繁玲, 2006)。自从Sprague等(1942)在玉米(Zea mays)杂交育种研究中首次提出2类配合力概念以来, 配合力分析在植物杂交育种中得到了普遍应用(张向群, 1987; 李周岐等, 2001), 并已成为评选优良亲本和杂种优势组合的重要依据(张爱民, 1994)。通过对杂交子代遗传分析, 一方面可选用强优势杂交组合建立双系杂交种子园或通过无性繁育加以固定利用, 另一方面可以选择一般配合力高的亲本作为新建无性系种子园的建园材料, 或在优良杂交组合中进行优良个体选择, 为高世代育种提供亲本材料。此外, 还可了解主要经济性状的基因作用方式, 为制订有效的遗传改良方案提供理论依据。
相对于欧洲赤松(Pinus sylvestris) (Blumenrother et al., 2001)、辐射松(P.radiata) (Jayawickrama et al., 2000)、火炬松(P.teada) (Mckeand et al., 1998)、杉木(Cunnighamia lanceolata) (王赵民等, 1988; 李力等, 2000)等, 马尾松(P.massoniana)的杂交育种起步较迟。目前本研究组已利用马尾松全双列(周志春等, 2004; 金国庆等, 2008a)、测交系(金国庆等, 2008b)等遗传交配设计子代测定林, 初步揭示了马尾松幼林生长和材性所受的遗传控制、杂种优势和近交衰退等规律, 初选出了一批优良杂交组合。巢式交配设计又称北卡罗来纳设计Ⅰ(NC designⅠ), 因供试母本个体只与1个父本个体杂交产生后代, 而父本个体却可以与若干母本个体杂交, 因此在动物遗传育种中应用较多(张帆等, 1996; 于飞等, 2008; 杨国梁等, 2008), 但在林木育种研究中相关研究报道则较少, 国内仅见梁一池(1999)对14年生杉木巢式交配子代生长性状的研究。本文利用设置在浙江省淳安县富溪林场12年生马尾松巢式遗传交配设计的子代测定林, 进一步开展其生长和木材基本密度的父本/母本效应和加性/显性基因效应等遗传分析, 揭示这些性状所受的遗传控制式样, 同时为生产和马尾松二代育种筛选出一批速生优质的杂交组合和个体。
1 材料与方法 1.1 试验材料1995年选用浙江省淳安县姥山林场马尾松一代育种群体中来自不同产地的12个和48个优树无性系分别做父本(主要来自广西和贵州)和母本(主要来自安徽、浙江、江西和福建), 按照巢式遗传交配设计进行控制授粉制种(图 1)。1996年11月收获试验种子, 1997年和1998年在淳安县富溪林场分别育苗和造林。子代测定林位于119° 03' E, 29°37' N, 海拔250~300 m, 年平均气温17 ℃, 年降雨量1 430 mm, 年日照1 951 h, 土壤为山地黄壤, 前茬为杉木人工林, 立地条件中等。试验采用完全随机区组设计, 5次重复, 8株小区, 株行距2 m×2 m, 带状整地。在育苗与造林时, 均以浙江省淳安县当地马尾松天然优良林分的种子为对照(CK)。

|
图 1 巢式交配 Figure 1 Pattern of nested mating design |
2009年12月在每重复的试验小区中选择3株生长最佳的植株, 测量其树高、胸径、树干通直度和树干圆满度(用树干下部2 m高处直径与胸径之比表示)等指标。树干通直度按通直、较通直、一般、弯曲、严重弯曲5级, 分别记为5, 4, 3, 2, 1。同时在植株上坡方位胸高处用6 mm直径的生长锥钻取一树皮至髓心的完整无疵木芯, 用最大含水量法(Smith, 1954)测定其木材基本密度(Di)。
1.3 统计分析单株材积按V=0.000062341803 ×D 1.31.8561497×H0.9568492估算(周志春等, 1993)。方差分析时树干通直度经平方根数据转换。以小区均值为单元, 采用SAS / STAT软件中的ANVOA进行性状方差分析以检验不同杂交组合间遗传差异的显著性(孔繁玲, 2006)。若杂交组合间差异显著, 按固定模型(ModelⅠ)估算父本效应和父本内母本效应, 按随机模型(Model Ⅱ)估算其遗传方差分量等(表 1) (沈熙环, 1990; 周志春等, 2004)。
|
|
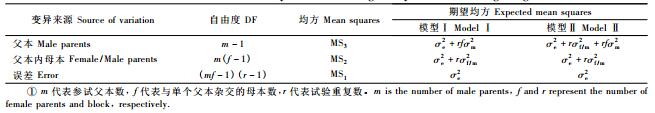
表 2方差分析结果表明, 除树干通直度在杂交组合间差异不显著外, 树高、胸径、单株材积、木材基本密度和树干圆满度等性状在不同杂交组合间的遗传差异皆达到1%或5%的显著水平, 意味着选用不同产地的无性系进行杂交可为马尾松高世代育种创造出变异丰富的基本育种群体, 也可为生产上创制和选育优良杂交组合和个体。进一步的统计分析结果显示, 马尾松生长性状和木材基本密度的父本/父本内母本效应均达到显著或极显著水平, 而树干圆满度仅其父本效应显著。
|
|
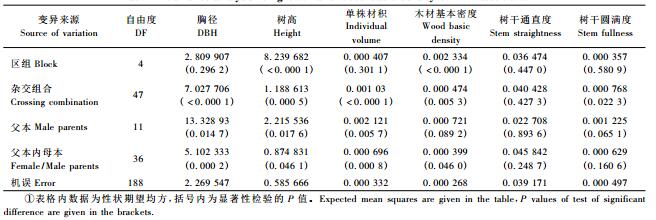
表 3为生长和木材基本密度的父本/母本效应(σ m 2/ σf/m2)和加性/显性基因效应(VA/VD)的方差分量大小, 用以说明其所受的遗传控制式样。结果发现, 胸径、树高、单株材积等生长性状及树干圆满度的父本/母本效应方差分量几乎相当, 说明父本和母本对子代的贡献率差异较小, 但木材基本密度则较多受母本效应的影响, 其母本效应是父本效应的1.56倍。根据遗传方差理论, 当近交系数等于0时, σm2=1 /4VA , σf/m2 = 1/4VA + 1/4VD, 据此估算加性(VA)和显性(VD)基因效应的方差分量(孔繁玲, 2006)。由表 3可见马尾松胸径和木材基本密度的加性基因效应分别是其显性基因效应的2.65倍和1.78倍, 说明基因作用的方式上二者以加性基因效应控制为主, 显性基因效应次之; 在对胸径和木材基本密度进行遗传改良时, 除重视亲本的一般配合力(GCA)选择外, 还应加强亲本间组配能力的筛选和测定。树高、单株材积和树干圆满度主要受加性基因效应控制, 显性基因效应很小, 可依据亲本表现对子代进行预测。
|
|
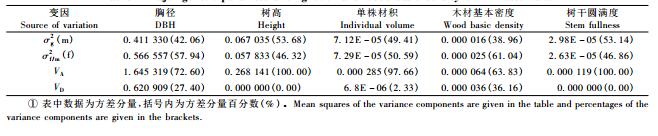
各父本无性系的胸径、树高、单株材积、木材基本密度和树干圆满度均值分别变化在11.74~14.76 cm, 9.67~10.83 m, 0.056~0.094 m3, 0.355~0.377 g·cm-3和0.913~0.944, 其GCA效应值列于表 4, 说明不同父本间的差异较大。在12个父本无性系中, 以南部广西产的1123无性系生长性状的GCA最大, 并显著高于其他11个父本无性系, 其胸径、树高和单株材积的GCA分别为2.255, 0.792和0.300, 福建产9134和湖南产7631无性系生长性状的GCA次之, 故以速生为目标时1123, 9134和7631是首选的优良父本; 福建产9515无性系生长最差, 其胸径、树高和单株材积的GCA最小且为负值, 分别为-0.765,-0.365和-0.009 1。比较木材基本密度和树干圆满度的GCA变化规律, 发现生长较快的父本无性系, 其木材基本密度不一定低, 树干圆满度不一定就好, 反之亦然。如福建产9515无性系, 不仅其生长性状的GCA最小, 而且木材基本密度的GCA也较低(-0.0027), 其树干圆满度的GCA在12个父本无性系中居于中等(-0.0026);湖南产7631不仅是速生的优良父本, 而且木材基本密度的GCA也较高, 达0.002 7, 但其树干圆满度的GCA却较小, 为-0.0046。
|
|
48个杂交组合的胸径、树高、单株材积、木材基本密度以及树干圆满度的变化范围分别为10.45~16.73 cm, 9.05~11.33 m, 0.043~0.123m3, 0.341~0.391 g·cm-3和0.877~0.951, 且其皆明显高于CK。对于胸径、树高和单株材积,5159×1123杂交组合表现最佳, 且明显高于其他杂交组合, 其值分别高于群体均值33.76%,12.83%和89.24%;5131×1123,178×7631, 5132×1123和5411×9134这4个杂交组合次之, 其值分别为13.67~15.03 cm, 10.66~11.16 m和0.080~0.093 m3; 9321×7631的胸径和单株材积在48个杂交组合中表现最差, 分别低于群体均值16.45%和33.68%。对木材基本密度的分析表明, 杂交组合的生长与木材密度的相关性较小。如6124×1312和5118×1108组合, 单株材积和木材基本密度均分别高于群体均值19.06%和12.48%以及4.96%和1.17%,说明这些杂交组合的木材基本密度不会因为速生而降低,在生产中对它们进行利用可同时达到对生长和材性的改良;而6128×1312和6302×1208杂交组合的单株材积和木材基本密度均分别低于群体均值14.63%和16.01%以及1.92%和1.24%, 在生长和材性上均属于较差的杂交组合。
马尾松主要用作纸浆纤维材和锯材。以木材干物质积累量(材积与木材基本密度的乘积)大于总体平均值10%、大于CK 70%为入选标准, 共选出10个纸浆纤维材优良杂交组合(表 5)。12年生时48个杂交组合干物质积累量变化在14.93~43.52 kg间, 而10个优良组合的干物质积累量在26.71~43.52 kg间, 较CK平均高出102.02%, 较总体平均值高出30.99%。作为优良锯材, 要求径生长量大、木材基本密度高、形质优。以胸径大于CK 20%, 木材基本密度和树干圆满度不低于CK为入选标准, 也选出10个锯材优良杂交组合, 其平均胸径13.53 cm, 较CK平均高出25.98%。
|
|
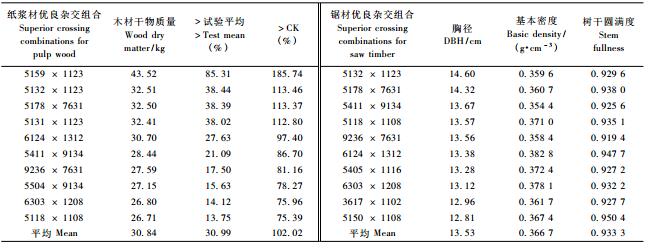
同样以纸浆纤维材和锯材为培育目标, 结合优良杂交组合开展优良个体选择以为二代育种增补亲本无性系, 每个组合至多只选择1株优株。若按单株干物质积累量大于群体平均值的100%、大于CK的200%为筛选标准, 共选出17株纸浆纤维材优株, 其平均单株干物质积累量58.13 kg, 较优良杂交组合高出88.49%。若按胸径大于群体平均值30%, 大于CK 60%, 木材基本密度和树干圆满度均大于对照和群体平均值作为选择标准, 共选出13株锯材优株, 其平均胸径为17.74 cm, 高出优良锯材杂交组合平均值的31.12%。在评选出的10个优良锯材杂交组合中, 有7个与纸浆纤维材优良组合相同, 而在选出的13个优良锯材单株中, 也有6个与纸浆纤维材优良单株相同。
3 结论与讨论12年生马尾松巢式交配设计的遗传测定结果表明, 除树干通直度外, 树高、胸径和单株材积、木材基本密度以及树干圆满度在不同杂交组合间存在显著的遗传差异, 如单株材积、木材基本密度和单株干物质积累量分别变化在0.043~0.123 m3, 0.341~0.391 g·cm-3和14.93~43.52 kg, 说明选用不同产地优树无性系开展杂交制种可为马尾松高世代育种创造变异丰富的基本群体。研究发现, 胸径、树高、单株材积及树干圆满度同时受父本和母本效应的控制, 且二者差异较小, 然而木材基本密度主要受母本效应的影响, 其母本效应是父本效应的1.56倍, 说明在对木材基本密度进行遗传改良时应重视对母本木材密度的选择。一般认为经过GCA测定的材料, 需进行SCA的选择和测定, 而未经过GCA测定与选择的材料, 则其GCA的选择比SCA更为重要(Sprague et al.,1942 )。本试验中因杂交亲本未经过GCA测定, 其生长和木材基本密度等主要经济性状的加性效应方差分量明显大于显性效应方差分量, 进一步支持了Sprague等(1942)的观点。分析表明, 12年生马尾松胸径和木材基本密度以加性效应控制为主, 显性效应次之, 与金国庆等(2008a)对马尾松测交系子代的研究结果一致, 但与周志春等(2004)和金国庆等(2008b)对马尾松全双列材料的研究结果相反, 推测可能与测定材料年龄不同或后者的杂交亲本是经GCA初步测定有关。与胸径和木材基本密度不同的是, 马尾松树高、单株材积和树干圆满度的遗传变异几乎完全来自于加性基因效应, 相比杉木(梁一池, 1999)和马尾松(金国庆等, 2008a)的研究结果, 本文得出的马尾松树高和单株材积的加性基因效应所占分量偏大。但李力等(2000)和张建章等(1998)分别对10.5和12年生杉木半双列遗传测定林的研究也发现树高和单株材积的加性基因效应方差几乎占总基因效应方差的100%。根据性状遗传控制大小及方式的研究结果, 对马尾松生长或木材密度进行遗传改良时, 可选择GCA高的亲本进行杂交以产生强优势杂种, 而对树干圆满度的改良, 选择干形优良的亲本即可。
王赵民等(1988)根据杉木生长性状配合力的研究结果认为, 利用GCA高的亲本进行杂交将会提高得到强优组合的机率。在本试验中未发现各杂交组合生长表现与父本无性系GCA间存在明显的线性相关性, 即由GCA高的父本无性系所组配的杂交组合其生长表现不一定优良, 如由单株材积GCA高的1123和7631父本无性系组配的杂交组合5159×1123和5178×7631, 其单株材积分别为0.123 m3和0.091 m3, 高出平均材积89.22%和40.22%, 而5129×1123和9321×7631组合的单株材积分别为0.057 m3和0.043 m3, 低于平均材积12.17%和33.74%, 推测可能与主要的生长性状如胸径生长和木材基本密度受母本效应影响较大有关。但这一结果并未否认王赵民等(1988)研究结论, 因亲本GCA包括父本和母本的GCA, 而到目前为止, 还未见合适的统计方法能对巢式交配设计中的母本GCA进行估算(王明庥, 2001)。梁一池(1999)利用杉木巢式交配子代测定材料估算了不同杂交组合生长性状的SCA效应值, 其估算方法存在一定的缺陷。另外, 在本文筛选的10个纸浆材优良杂交组合中, 以干物质积累量GCA最高的3个父本无性系1123, 9134和7631所组配的优良杂交组合就有7个, 占入选优良杂交组合数70%, 这进一步说明了GCA育种的重要性。在生产实践中, 若要获得强优势杂交组合, 不仅要求父本无性系的GCA高, 同时也要求母本无性系具有高的GCA。以干物质积累量和胸径等为选择指标, 分别选出纸浆纤维材和锯材各10个优良杂交组合。在选择的10个纸浆纤维材优良组合中, 以5159×1123组合的干物质积累量最高, 分别高出总体平均值和CK 85.31%和185.74%, 5178×7631, 5132×1123, 5131×1123, 6124×1312和5411×9134这5个杂交组合的干物质积累量次之, 分别高出总体平均值和CK 21.09%~38.44%和86.7%~113.46%, 可通过有性制种无性繁育的途径加以利用。此外, 还分别选择出了17个和13个纸浆纤维材和锯材为培育目标的优良单株。
金国庆, 秦国峰, 储德裕, 等. 2008a. 马尾松测交系杂交子代生长性状遗传分析[J]. 林业科学, 44(1): 70-76. |
金国庆, 秦国峰, 周志春, 等. 2008b. 马尾松生长性状的交配效应的遗传分析及杂交组合选择[J]. 林业科学, 44(6): 28-33. |
孔繁玲. 2006. 植物数量遗传学[M]. 北京: 中国农业出版社: 115-285.
|
李力, 施季森, 陈孝丑, 等. 2000. 杉木两水平双列杂交亲本配合力分析[J]. 南京林业大学学报, 24(5): 9-13. |
李周岐, 王章荣. 2001. 林木杂交育种研究新进展[J]. 西北林学院学报, 16(4): 93-96. |
梁一池. 1999. 杉木配合力第一轮回的选择: Ⅰ遗传参数估计及配合力效应分析[J]. 福建林学院学报, 19(3): 193-196. |
马常耕. 1995. 我国杨树杂交育种的现状与发展对策[J]. 林业科学, 31(1): 60-66. |
沈熙环. 1990. 林木育种学[M]. 北京: 中国林业出版社: 135-152.
|
王赵民, 陈益泰. 1988. 杉木主要生长性状配合力分析及杂种优势的利用[J]. 林业科学研究, 1(6): 614-623. |
王明庥. 2001. 林木遗传育种学[M]. 北京: 中国农业出版社: 166-171.
|
杨国梁, 王军毅, 孔杰, 等. 2008. 罗氏沼虾大规模家系构建与培育技术研究[J]. 海洋水产研究, 29(3): 62-66. |
于飞, 张庆文, 孔杰, 等. 2008. 大菱鲆不同进口群体杂交后代的早期生长差异[J]. 水产学报, 32(1): 58-64. |
张爱民. 1994. 植物育种亲本选配的理论和方法[M]. 北京: 中国农业出版社: 1-12.
|
张帆, 毛盛贤. 1996. 乌鸡的性状遗传率、遗传相关和杂种优势[J]. 首都师范大学学报, 17(2): 79-83. |
张建章, 游昌顺. 1998. 杉木半双列杂交后代生长性状的配合力分析[J]. 浙江林学院学报, 15(2): 138-144. |
张向群. 1987. 玉米自交系2种配合力在杂种一代的表现[J]. 作物学报, 13(2): 135-142. |
周志春, 傅玉狮, 吴天林. 1993. 马尾松生长和材性的地理遗传变异及最优种源区的划定[J]. 林业科学研究, 6(5): 556-564. |
周志春, 金国庆, 秦国峰, 等. 2004. 马尾松纸浆材重要经济性状配合力及杂种优势分析[J]. 林业科学, 40(4): 52-57. DOI:10.11707/j.1001-7488.20040409 |
Blumenrother M, Bachmann M, Muller-Starck G. 2001. Genetic characters and diameter growth of provenances of Scots Pine (Pinus sylvestris L.)[J]. Silvae Genetica, 50(5 /6): 212-222. |
Jayawickrama K J S, Carson M J. 2000. A breeding strategy for the New Zealand radiata pine breeding cooperative[J]. Silvae Genetica, 49(2): 82-90. |
King J N, Carson M J, Johnson G R. 1997. Analysis of disconnected diallel mating designs Ⅰ: Results from a third generation progeny test of the New Zealand radiata pine improvement programme[J]. Silvae Genetica, 47(2 /3): 80-87. |
Mckeand S E, Bridgwater F E. 1998. A strategy for the third breeding cycle of loblolly pine in the southeastern U.S[J]. Silvae Genetica, 47(4): 223-235. |
Smith D M. 1954. Maximum moisture content method for determining specific gravity of small wood samples. US Forest Service, Forest Products Laboratory, Report No: 2014.
|
Sprague G F, Tatum L A. 1942. General VS specific combining ability in single crosses of corn[J]. J Am Soc Agrom, 34(4): 923-932. |