 2011, Vol. 47
2011, Vol. 47文章信息
- 刘建锋, 肖文发, 郭明春, 吴焕萍, 江泽平
- Liu Jianfeng, Xiao Wenfa, Guo Mingchun, Wu Huanping, Jiang Zeping
- 基于3-PGS模型的中国陆地植被NPP格局
- Pattern Analysis of Net Primary Productivity of China Terrestrial Vegetation Using 3-PGS Model
- 林业科学, 2011, 47(5): 16-22.
- Scientia Silvae Sinicae, 2011, 47(5): 16-22.
-
文章历史
- 收稿日期:2010-07-21
- 修回日期:2011-04-07
-
作者相关文章



2. 中国林业科学研究院森林生态环境与保护研究所 北京 100091;
3. 河北科技师范学院园 艺科技学院 秦皇岛 066004;
4. 国家气象中心 北京 100081
2. Research Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091;
3. College of Horticulture Science and Technology, Heibei Normal University of Science and Technology Qinhuangdao 066004;
4. National Meteorological Center Beijing 100081
植被净初级生产力(net primary productivity,简称NPP)作为地表碳循环的重要组成部分,直接反映植被群落在自然环境条件下的生产能力,表征陆地生态系统的质量状况,是判定生态系统碳汇和调节生态过程的主要因子(Field et al., 1998),在全球变化及碳平衡中扮演着重要角色。传统的基于样点数据估算NPP方法易受空间尺度限制,因此诞生了许多模型估算NPP的方法,可归结为以下3类: 1)气候估算模型代表模型有Miami模型(Lieth,1973)、Thornthwaite纪念模型(Lieth et al., 1972)、Chikugo模型(Uchijima,1985)和周广胜综合模型(周广胜等,1995)等; 2)生理生态过程模型代表模型有BIOME-BGC模型(Running et al., 1993)、CENTURY模型(Parton et al., 1993)和TEM模型(Raich et al., 1991)等; 3)光能利用效率模型代表模型有CASA模型(Potter et al., 1993)、SDBM模型(Knorr et al., 1995)、GLO-PEM模型(Prince et al., 1995)和3-PGS模型(Coops et al., 1998)等。以上3类模型都各有优缺点,如气候模型简单但估算的是潜在的NPP,过程模型估算较准确但所需参数太多且难以获取,光能利用效率模型适合较大空间尺度NPP估算,但机理不清晰(朱文泉等,2005)。
陆地植被NPP具有明显的时空变化特征,空间变化主要与区域环境条件、气候特征、植被类型有关; 而时间动态表现为季节和年际变化(陶波等,2003)。随着遥感技术的发展,区域或全球长时序的植被动态观测已成为可能,而且作为光能利用效率模型核心的冠层光合有效辐射利用分量亦可由遥感手段获得,从而加速了光能利用效率模型在区域或全球NPP估算研究中的应用。3-PGS模型由于模型不需考虑地下生态过程,且仅以固定比值(NPP/GPP=0.47±0.05)来估算生态系统的呼吸消耗,这使得估算过程相对便捷(Landsberg et al., 1997; Coops et al., 1998)。目前该模型已被广泛用于森林生产力估算(Coops et al., 2001a; 2001b; 2001c; 2007; 2009; Tickle et al., 2001; Nightingale et al., 2007)。该模型的应用结果表明,3-PGS是估算区域植被NPP或GPP的有效手段之一。如Coops等(2001a; 2001c)利用3-PGS模型估算美国俄勒冈州森林NPP,其估算结果与BIOME-BGC模型估算结果和实测结果显著相关(R2分别为0.85和0.75)。Nightingale等(2007)及Coops等(2009)利用3-PGS模型估算美国森林GPP,发现估算结果与全球MODIS标准GPP产品(MOD-17)估算结果具有较好的一致性。Coops等(2007)应用3-PGS对道格拉斯冷杉(Pseudotsuga menziesii)林GPP的估算结果与涡度相关方法得到的结果显著相关(R2=0.86)。我国已有部分学者应用其他光能利用效率模型对全国植被NPP进行估算(朴世龙等,2001a; 2001b; 孙睿等,2001; 陈利军等,2002; 朱文泉等,2007; 侯英雨等,2007),但未见3-PGS模型的国内应用案例。本研究应用3-PGS模型估算我国植被NPP总量并分析其时空格局(2003—2007年),为我国植被碳汇估算提供参考。
1 3-PGS模型简介3-PGS模型是Coops等(1998)在生理过程模型3-PG(Landsberg et al., 1997)基础上,应用空间遥感数据发展形成的。和其他光能利用效率模型一样,该模型利用植被所吸收的光合有效辐射与光能利用效率来计算植被NPP,考虑到了温度、霜冻、水汽压亏缺和土壤有效水分等环境因子对植被光能利用效率的影响。该模型可表达为NPP=Cpp×FPAR×PAR×ε0×; [fT×fF×Min(fD,fθ)],式中: Cpp为植被总初级生产(GPP)转化为NPP的系数,模型默认为0.47;FPAR为光合有效辐射吸收分量; PAR为光合有效辐射,默认为太阳总辐射值的45%;ε0为植被最大光能利用效率,默认值为1.80 g C·MJ-1。fT,fF,fD和fθ分别为气温、霜冻日数、水汽压亏缺和土壤有效水分含量的函数,每个函数描述一种限制因子,其值在0和1之间。Min(fD,fθ)表示取fD与fθ间最小值。
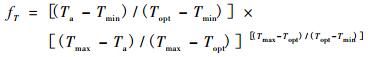
|
式中: Ta为月平均气温(℃); Tmax和Tmin分别为月平均最高和最低气温(℃); Topt为植被最适生长气温(℃)。
fF=1-fn/d,fn=-aTmin+b,式中: d为每月天数,a和b为经验系数; 如果fn<0,则fn取0。
fD=exp(-kD),式中: k为常数,D为水汽压亏缺。
fθ=1/{1+[(1-rθ)/cθ] n(θ)},rθ=[θ+(P-ET)]/θm(如分子大于分母,表明土壤水分吸持达到饱和而形成径流,则rθ取θm; 如果分子是负值,则rθ为0),式中: cθ和n(θ)为土壤水分特征参数; θ为当月土壤含水量,θm为土壤最大可利用水分含量; P为月降水量(mm); ET为月蒸散量(mm),根据彭曼公式求算。
2 数据来源与处理收集全国685个标准气象台站观测统计的2003—2007年逐月的平均气温、最低气温和降水量数据,以及95个地面辐射台站的太阳辐射数据。利用ARCGIS(V9.2)的插值模块进行空间插值(克里格法),生成地面分辨率为1 km×1 km的栅格图层。
土壤质地数据自世界和谐土壤数据库http://www.iiasa.ac.at提取(中国土壤质地原始数据由中国科学院南京土壤研究所汇交,分辨率为1 km×1 km)。
2003—2007年逐月的FPAR数据来源于美国波士顿大学气候与植被研究组(http://cybele.bu.edu/modismisr/index.html)。利用ENVI(V4.2)进行拼接、投影变换和裁切,分辨率为1 km×1 km。该数据集是利用中分辨率成像光谱仪(moderateresolution imaging spectroradiometer,简称MODIS)数据按辐射传输模型运算加工而成,在全球植被参数与NPP/GPP估算中应用广泛。
全国气候区划是根据国家气象局1981年版的中国气候区划图进行矢量化获得的,以字母A,B,C,D,E,F,G,H和J分别代表寒温带、中温带、暖温带、北亚热带、中亚热带、南亚热带、北热带、中热带和高原气候区,藏东南地区气候特殊,标示为JEFG。植被覆盖类型采用MODIS-12(UMD)分类系统,共12类,依次为常绿针叶林、常绿阔叶林、落叶针叶林、落叶阔叶林、混交林、密闭灌丛、稀疏灌丛、稀树草原、草原、草地、农地和荒漠,分别表示为ENF,EBF,DNF,DBF,MF,CS,OS,WS,SA,GL,CL和DE。
3 结果与分析 3.1 中国陆地植被NPP空间格局利用3-PGS估算结果表明,2003—2007年中国陆地植被NPP年均值为315.99 gC·m-2 a-1,NPP年均总量为2.98 PgC,其中森林植被占40.77%。从图 1可以看出,我国陆地植被NPP分布主要受水热条件限制,从东南沿海向西北部逐渐减小,西北沙漠地区最小,其主要原因是西部地区植被稀少,气候干旱,而东部地区气候湿润,水热条件好,能更好地满足植被生长(陶波等,2003)。

|
图 1 2003—2007年全国年均NPP分布 Figure 1 Distribution of annual mean NPP in China from 2003 to 2007 |
从气候带分布来看(图 2),以中热带(H,海南岛中南部)最高(1 122.88 gC·m-2 a-1),北热带(G,云南南部、雷州半岛、台湾南部)次之(9 74.03 gC·m-2 a-1),南亚热带(F,云南中南部、广西南部、广东中南部和福建南部地区)又次之(813.31 gC·m-2 a-1)。年均NPP最低值发生在高原气候区(J,青藏高原)仅为91.69 gC·m-2 a-1,然后是中温带(B)194.56 gC·m-2 a-1。藏东南地区(JEFG)受印度洋季风影响,水热丰富,具有热带、亚热带典型海洋性山地气候(张新时,1978),年均NPP水平与北亚热带和中亚热带接近,达到626.72 gC·m-2 a-1。不同气候带NPP年均总量方面,则以中亚热带最高(E,920.90 TgC),中热带最低(H,5.53 TgC),这与面积有关。

|
图 2 不同气候带NPP年均值及年总量 Figure 2 Mean and sum of annual NPP over various climate types A:寒温带North temperate; B:中温带Middle temperate; C:暖温带South temperate; D:北亚热带North subtropical; E:中亚热带Middle subtropical; F:南亚热带South subtropical; G:北热带North tropical; H:中热带Middle tropical; J:高原气候区Plataeu climate; JEFG:藏南气候区South Tibet climate. |
由图 3可见,我国主要植被类型的年均NPP超过了400 gC·m-2 a-1,其中常绿阔叶林年均NPP最高,达到1 066.17 gC·m-2 a-1,然后依次为混交林(731.27 gC·m-2 a-1)、常绿针叶林(555.87gC·m-2 a-1)、落叶阔叶林(656.32 gC·m-2 a-1)和落叶针叶林(446.12 gC·m-2 a-1),荒漠年均NPP最低,仅为4.36 gC·m-2 a-1。全国不同植被的NPP年均总量方面,以农地最高(972.82 TgC),荒漠最低(4.37 TgC)。

|
图 3 不同植被类型NPP年均值及年总量 Figure 3 Mean and sum of annual NPP over various vegetation types ENF:常绿针叶林Evergreen needle forest; EBF:常绿阔叶林Evergreen broadleaf forest; DNF:落叶针叶林Deciduous needle fo rest; DBF:落叶阔叶林Deciduous broadleaf forest; MF:混交林Mixed forest; CS:郁闭灌丛Closed shrubland; OS:稀疏灌丛Open shrubland; WS:稀树草原Woody savannas,; SA:草原Savannas; GL:草地Grassland; CL:农地Cropland; DE:荒漠Desert |
图 4为2003—2007年NPP月平均值,由图可见,我国陆地植被NPP的季节变化明显。在夏季7和8月全国各地区太阳辐射丰富,水热条件适合植物生长,NPP月均值达到峰值,分别为79.70和75.54 g C·m-2。1和12月是我国平均太阳辐射和气温最低的月份,秦岭以北地区的植被在这段时间内基本停止生长,而以南地区植被受温度等环境因素的影响,光能转化效率比夏季低(朴世龙等,2001a)。12月份的NPP均值下降至全年最低值(1.12 gC·m-2)。

|
图 4 中国植被NPP季节动态(2003—2007) Figure 4 Seasonal dynamic of NPP in China vegetation(2003—2007) |
从表 1可以看出,北热带和中热带植被在5月NPP达到峰值,南亚热带在9月达到峰值,其他气候带均在7,8月出现峰值。寒温带7月植被NPP最高,为176.20 gC·m-2,高原气候区植被峰值仅为33.04gC·m-2。寒温带、中温带、暖温带和高原气候区植被光合生产集中在5—9月; 而亚热带和热带地区全年水热较丰富,植被整年均有物质生产与积累。
|
|

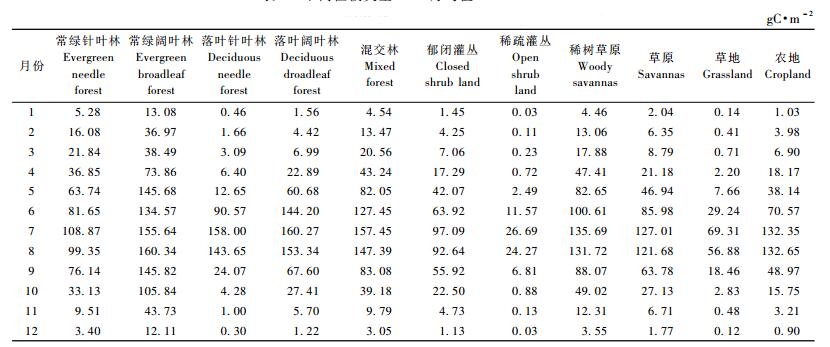
我国不同植被类型的NPP月均值变化规律明显(表 2),峰值均出现在7,8月,峰值较高的植被类型主要是森林,以常绿阔叶林最高,达160.34gC·m-2,峰值较低的植被是草地和稀疏灌丛,分别为69.31和26.69 gC·m-2。
|
|
2003—2007年我国陆地植被NPP年均总量为2.98 PgC,NPP年均值约为315.99 gC·m-2 a-1。与其他应用光能利用效率模型的研究结果相比,本研究对NPP年均总量的模拟结果属中等水平(2.98 PgC),高于朴世龙等(2001a; 2001b)利用CASA模型估算得到的1.95和1.80 PgC,略高于孙睿(2000)估算得到的2.645 PgC,低于陈利军等(2002)估算得到的6.13 PgC和陈斌等(2007)利用C-Fix模型得到的4.37 PgC,略低于朱文泉等(2007)估算得到的3.12 PgC。与生理过程模型相比,接近陶波等(2003)利用CEVSA模型估算得到的3.09 PgC,高于冯险峰(2004)利用BEPS-China模型得到的2.24 PgC,而低于Xiao(1998)利用TEM模型得到的3.653 PgC。高志强等(2008)利用3个光能利用效率模型(CASA,GLOPEM和GEOLUE)和2个生理过程模型(CEVSA和GEOPRO)估算中国植被NPP年总量约为2.864±1 PgC。
NPP的基本分布趋势与众多模型应用结果相类似,即随水热条件梯度变化,从西到东,从北到南,NPP逐渐递增。热带植被NPP水平最高; 而高原气候区寒冷干旱,NPP最低。降水量是影响我国植被NPP的主要因子(周广胜等,1998; 朴世龙等,2001b; 陶波等,2003;侯雨英等,2007)。本研究模型估算结果与Cramer等(1999)对全球不同植被类型NPP预测的范围一致,即最高的NPP出现在水热条件良好的热带地区(>1 000 g C·m-2 a-1),温带地区的NPP居中(500~700 g C·m-2 a-1),NPP最低值出现在寒冷和干旱地区(<200g C·m-2 a-1)。不同植被类型NPP以常绿阔叶林最高,而荒漠最低,与朴世龙等(2001b)的研究结果一致。将本研究基于3-PGS模型模拟的主要森林植被类型NPP结果与其他部分研究结果和实测值比较(孙睿,2000; 朱文泉等,2007; 朴世龙等,2001a; 陶波等,2003; 刘世荣等,1997),结果表明,3-PGS模型的估算结果基本落在实测值范围之内,也基本处在其他模型估算的最大值和最小值之间,说明本模型估算结果还是比较准确的。在NPP季节动态方面,不同气候类型和植被类型基本在7或8月达到峰值,峰值高的类型主要是热带亚热带的森林植被,与侯英雨等(2007)研究结果相似。
综上所述,与其他模型相比,3-PGS模型对我国陆地植被NPP总量与时空格局的模拟具有一定的可靠性,能够反映出我国陆地植被NPP的时间动态和分布格局。然而,3-PGS模型估算过程的任一变量都有可能是NPP估算的误差来源,诸如模型应用1.80 gC·MJ-1为植被的最大光能利用效率颇有争议,该值会因植被类型不同而有所不同(Ruimy et al.,1994;彭少麟等,2000; 朱文泉等,2006; Waring et al., 2010); 其次是NPP/GPP比值也会因植被和环境条件不同而存在差异(Waring et al., 1998; Zhang et al., 2009)。太阳辐射、温度和土壤水分(降水)等栅格运算图层则会因对应站点的数量和分布均匀程度以及插值方法而产生一定的偏差。这几个方面也是应用或提高3-PGS及类似模型的模拟精度首先需要考虑的。另外,模型关键参数的验证与敏感性分析(如最大光能利用效率、土壤水分等)是获得准确估算结果的重要步骤; 而利用模型分析植被NPP对气候变化的响应是研究未来气候背景下植被碳汇潜力的重要方法。这些都将是下一步需要深入探讨的问题。
陈斌, 王绍强, 刘荣高, 等. 2007. 中国陆地生态系统NPP模拟及空间格局分析[J]. 资源科学, 29(6): 45-53. |
陈利军, 刘高焕, 励惠国. 2002. 中国陆地植被净第一性生产力遥感动态监测[J]. 遥感学报, 6(2): 129-135. DOI:10.11834/jrs.20020210 |
冯险峰. 2004. 基于过程的中国陆地生态系统生产力和蒸散遥感研究[J]. 中国科学院地理科学与资源研究所博士学位论文. |
高志强, 刘纪远. 2008. 中国植被净初级生产力的比较研究[J]. 科学通报, 53(3): 317-326. |
侯英雨, 柳钦火, 延昊, 等. 2007. 我国陆地植被净初级生产力变化规律及其对气候的响应[J]. 应用生态学报, 18(7): 1546-1553. |
刘世荣, 徐德应, 王兵, 等. 1997. 气候变化对中国森林生产力的影响//徐德应.气候变化对中国森林影响研究[M]. 北京: 中国科技出版社: 75-93.
|
彭少麟, 郭志华, 王伯荪. 2000. 利用GIS和RS估算广东植被光利用率[J]. 生态学报, 20(6): 903-909. |
朴世龙, 方精云, 郭庆华. 2001a. 利用CASA模型估算我国植被净第一性生产力[J]. 植物生态学报, 25(5): 603-608. |
朴世龙, 方精云, 郭庆华. 2001b. 1982—1999年我国植被净第一性生产力及其时空变换[J]. 北京大学学报:自然科学版, 37(4): 563-569. |
孙睿, 朱启疆. 2000. 中国陆地植被净第一性生产力及季节变化研究[J]. 地理学报, 55(1): 36-45. DOI:10.11821/xb200001004 |
陶波, 李克让, 邵雪梅, 等. 2003. 中国陆地净初级生产力时空特征模拟[J]. 地理学报, 58(3): 372-380. DOI:10.11821/xb200303006 |
张新时. 1978. 西藏植被的高原地带性[J]. 植物学报, 20(2): 140-149. |
周广胜, 张新时. 1995. 自然植被的净第一性生产力模型初探[J]. 植物生态学报, 17(3): 1-8. |
周广胜, 郑元润, 陈四清, 等. 1998. 自然植被净第一性生产力模型及其应用[J]. 林业科学, 34(5): 2-10. |
朱文泉, 陈云浩, 徐丹, 等. 2005. 陆地植被净初级生产力计算模型研究进展[J]. 生态学杂志, 24(3): 296-300. |
朱文泉, 潘耀忠, 何浩, 等. 2006. 中国典型植被最大光利用率模拟[J]. 科学通报, 51(6): 700-706. |
朱文泉, 潘耀忠, 张锦水. 2007. 中国陆地植被净初级生产力遥感估算[J]. 植物生态学报, 31(3): 413-424. |
Coops N C, Black T A, Jassal R S, et al. 2007. Comparison of MODIS, eddy covariance determined and physiologically modeled gross primary production(GPP)in a Douglas-fir forest stand[J]. Remote Sensing and Environment, 107(3): 385-401. DOI:10.1016/j.rse.2006.09.010 |
Coops N C, Ferster C J, Waring R H, et al. 2009. Comparison of three models for predicting gross primary production across and within fo rested ecoregions in the contiguous United States[J]. Remote Sensing of Environment, 113(3): 680-690. DOI:10.1016/j.rse.2008.11.013 |
Coops N C, Waring R H, Landsberg L L. 1998. Assessing forest productivity in Australia and New Zeal and using a physiologicallybased model driven with averaged monthly weather data and satellite derived estimates of canopy photosynthetic capacity[J]. Forest Ecology and Management, 104(1-3): 113-127. DOI:10.1016/S0378-1127(97)00248-X |
Coops N C, Waring R H, Brown S R, et al. 2001a. Comparisons of predictions of net primary production and seasonal patterns in water use derived with two forest growth models in Southwestern Oregon[J]. Ecological Modelling, 142(1-2): 61-81. DOI:10.1016/S0304-3800(01)00279-4 |
Coops N C, Waring R H. 2001b. Estimating forest productivity in the eastern Siskiyou Mountains of southwestern Oregon using a satellite driven process model, 3-PGS[J]. Canadian Journal of Forest Research, 31(1): 143-154. DOI:10.1139/x00-146 |
Coops N C, Waring R H. 2001c. The use of multiscale remote sensing imagery to derive regional estimates of forest growth capacity using 3-PGS[J]. Remote Sensing and Environment, 75(3): 324-334. DOI:10.1016/S0034-4257(00)00176-0 |
Cramer W, Kicklighter D Q, Bondeau A, et al. 1999. Comparing global models of terrestrial net primary productivity(NPP): overview and key results[J]. Global Change Biology, 5(Suppl.1): 1-15. |
Field C B, Behrenfeld M J, Randerson J T, et al. 1998. Primary production of the biosphere: integrating terrestrial and oceanic components[J]. Science, 281(5374): 237-240. DOI:10.1126/science.281.5374.237 |
Knorr W, Heimann M. 1995. Impact of drought stress and other factors on seasonal land biosphere CO2 exchange studies through an atmospheric tracer transport model[J]. Tellus B, 47(4): 471-489. DOI:10.3402/tellusb.v47i4.16062 |
Landsberg J J, Waring R H. 1997. A generalized model of forest productivity using simplified concepts of radiation-use efficiency, carbon balance, and partitioning[J]. Forest Ecology and Management, 95(3): 209-228. DOI:10.1016/S0378-1127(97)00026-1 |
Lieth H, Box E O. 1972. Evapotranspiration and primary productivity, CW Thornthwaite Memorial Model[J]. Publications in Climatology, 25(2): 37-46. |
Lieth H. 1973. Primary production: terrestrial ecosystems[J]. Human Ecology, 1(4): 303-332. DOI:10.1007/BF01536729 |
Nightingale J M, Coops N C, Waring R H, et al. 2007. Comparison of MODIS gross primary production estimates for forests across the U.S.A.with those generated by a simple process model, 3-PGS[J]. Remote Sensing of Environment, 109(4): 500-509. DOI:10.1016/j.rse.2007.02.004 |
Parton W J, Scurlock J M O, Ojima D S, et al. 1993. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide[J]. Global Biogeochemical Cycles, 7(4): 785-809. DOI:10.1029/93GB02042 |
Potter C S, Randerson J T, Field C B, et al. 1993. Terrestrial ecosystem production: a process model based on global satellite and surface data[J]. Global Biogeochemical Cycles, 7(4): 811-841. DOI:10.1029/93GB02725 |
Prince S D, Goward S N. 1995. Global net primary production: a remote sensing approach[J]. Journal of Biogeography, 22(4/5): 815-835. DOI:10.2307/2845983 |
Raich J W, Rastetter E B, Melillo J M, et al. 1991. Potential net primary productivity in south America: application of a global model[J]. Ecological Application, 1(4): 399-429. DOI:10.2307/1941899 |
Ruimy A, Saugier B. 1994. Methodology for the estimation of terrestrial net primary production from remotely sensed data[J]. Journal of Geophysical Research, 97(D3): 5263-5284. |
Running S W, Hunt E R. 1993. Generalization of a forest ecosystem process model for other biomes, BIOME-BGC, and an application fo r global-scale models//Ehleringer J R, Field C B.Scaling physiological processes: leaf to globe[J]. New York: Academic Press Inc: 141-158. |
Tickle P K, Coops N C, Hafner S D, et al. 2001. Assessing forest productivity at local scales across a native eucalypt forest using a process model, 3PG-SPATIAL[J]. Forest Ecology and Management, 152(1-3): 275-291. DOI:10.1016/S0378-1127(00)00609-5 |
Uchijima Z. 1985. Agroclimate evaluation of net primary productivity of natural vegetation(1)Chikugo model for evaluating net primary productivity[J]. Journal of Agricultural Meteorology, 40(4): 343-352. DOI:10.2480/agrmet.40.343 |
Waring R H, Coops N C, Landsberg J J. 2010. Improving predictions of fo rest growth using the 3-PGS model with observation made by remote sensing[J]. Forest Ecology and Management, 259(9): 1722-1729. DOI:10.1016/j.foreco.2009.05.036 |
Xiao X M. 1998. Net primary production of terrestrial ecosystems in China and its equilibrium responses to changes in climate and atmospheric CO2 concentration[J]. Acta Phytoecologica Sinica, 22(2): 97-118. |
Zhang Y J, Xu M, Chen H, et al. 2009. Global pattern of NPP to GPP ratio derived from MODIS data: effects of ecosystem type, geographical location and climate[J]. Global Ecology and Biogeography, 18(3): 280-290. DOI:10.1111/geb.2009.18.issue-3 |