 2011, Vol. 47
2011, Vol. 47文章信息
- 王晓阳, 李火根
- Wang Xiaoyang, Li Huogen
- 鹅掌楸苗期生长杂种优势的SSR分析
- Possible Mechanism Analysis for Heterosis of Hybrid Liriodendron Based on Seedling Growth and SSR Markers
- 林业科学, 2011, 47(4): 57-62.
- Scientia Silvae Sinicae, 2011, 47(4): 57-62.
-
文章历史
- 收稿日期:2010-01-15
- 修回日期:2010-04-20
-
作者相关文章



2. 中国热带农业科学院香料饮料研究所 万宁 571533
2. Spice and Beverage Crops Research Institute, Chinese Academy of Tropical Agricultural Science Wanning 571533
植物杂种优势的利用在植物生产中发挥了非常重要的作用, 取得了巨大的经济效益。相对于杂种优势的利用, 杂种优势的基础理论研究远远滞后, 严重制约着杂种优势的深入探索和进一步利用, 揭示杂种优势遗传基础成为植物遗传学领域中一个亟待解决的难题(兰进好等, 2005)。迄今为止, 许多学者利用分子标记对亲本遗传差异与杂种优势间关系进行了探索, 但研究结果不尽一致。一些研究认为, 亲本间的遗传距离与杂种优势间相关显著, 可作为选择杂交亲本的一项参考指标(许占友, 1997; 史延丽等, 2008; Stuber et al., 1992; Smith et al., 1990; Lee et al., 1989)。但也有研究表明, 亲本间的分子遗传差异与杂种优势相关程度较低, 不能应用于杂交亲本选配(刘宏伟等, 2005; Dudley et al., 1991; Martin et al., 1995; Barbosa et al., 1996)。
杂交鹅掌楸(Liriodendron chinense × tulipifera)为鹅掌楸(L.chinense)与北美鹅掌楸(L.tulipifera)的人工杂种, 具有非常明显的杂种优势, 其生长快, 适应能力强, 树形美观, 花色鲜艳, 无性繁殖容易, 因而具有极广阔的开发利用前景(王章荣, 1997)。对于鹅掌楸杂种优势的遗传基础, 虽然国内学者通过生长、光合效率等表型, 以及RAPD分子标记进行过探索(叶金山等, 2002; 李周岐等, 2002), 但均存在一定的局限性, 不能给出令人信服的解释。
本研究以12个2年生鹅掌楸种间杂交组合子代及其亲本自由授粉子代苗木为材料, 利用SSR分子标记分析苗期生长杂种优势与亲本遗传距离及杂种F1杂合度的相关性, 期望为鹅掌楸杂种优势遗传基础研究提供参考。
1 材料与方法 1.1 试验材料2005年5月, 选取成年的鹅掌楸6株(WYS, H, S, C5, J, J1)、北美鹅掌楸5株(ZZY, BK1, L, M, BM)作为杂交亲本, 进行人工控制授粉, 共得到12个种间杂交组合(BM×C5, ZZY×WYS, BK1×H, H×L, BK1×S, S×BK1, M×S, S×M, BM×S, S×BM, L×J1, J×L)。2005年11月初采种, 同时采集上述9个亲本的自由授粉种子(H和C5未采集到自由授粉种子)。2006年1月播种于南京林业大学校园苗圃。2007年3月在镇江扬子江林业有限公司苗圃进行苗期试验。采用完全随机区组设计, 4次重复, 10株小区, 株行距40 cm×40 cm。2007年11月生长季结束后, 对参试的12个杂交组合子代及9个交配亲本的自由授粉子代逐株调查苗高、地径。2007年7月中旬采集上述11个亲本的无病斑嫩叶用于亲本遗传距离分析, 采集9个杂交组合(BM×C5, BK1×S, S×BK1, M×S, S×M, BM×S, S×BM, L×J1, J×L)的无病斑嫩叶用于子代群体杂合度分析, 每个组合采样40株。
1.2 杂种优势分析杂种优势计算公式(卢庆善等, 2002):超中亲优势Hm=(F1-MP)/MP×100%;超高亲优势Hh=(F1-HP)/HP×100%。式中, F1为杂种子代的性状平均值, MP为2个亲本性状的平均值, HP为双亲中较高亲本的性状值。由于各交配亲本树龄达12年, 不能直接以其性状值为对照来估算杂种优势, 因此, 以各亲本的自由授粉子代性状平均值作为该亲本的性状值。
由于2个鹅掌楸亲本(H和C5)未采集到自由授粉种子, 导致无法估算该2个亲本交配组合子代(BK1×H, H×L, BM×C5)的超中亲优势。鉴于以北美鹅掌楸为母本的自由授粉子代平均生长量大于以鹅掌楸为母本的自由授粉子代平均生长量(张晓飞, 2008), 因此, 分别以BK1, L, BM 3个亲本的自由授粉子代的平均生长量作为参照, 计算上述3个杂交组合子代的超高亲优势。
1.3 DNA提取基因组DNA的提取采用改进的CTAB裂解-硅珠吸附法(张博等, 2004)。
1.4 SSR分析SSR引物来自作者所在实验室自主开发的鹅掌楸属特异性EST-SSR引物(Xu et al., 2006)。PCR总反应体系为10 μL, 包括1×buffer, 2.5 mmol·L-1 MgCl2, 0.2 mmol·L-1 dNTPs, 0.25 μmol·L-1 SSR引物, 0.25 U Taq酶, 20~40 ng模板DNA。扩增反应程序采用Touch-down PCR:94 ℃预变性4 min, 随后15个Touch-down循环(94 ℃变性15 s, 60 ℃退火15 s, 72 ℃延伸30 s, 每次循环的退火温度降0.7 ℃), 再进行一般性PCR扩增, 共15个循环, 每个循环包括94 ℃ 15 s, 49.5 ℃ 15 s, 72 ℃ 30 s, 最后于72 ℃延伸20 min。扩增产物用8%聚丙烯酰胺变性凝胶电泳分离检测, 银染法显带分析。
1.5 亲本遗传距离与子代杂合度分析将每一个SSR扩增位点看成是1个基因位点, 扩增位点中的每一条带看成是1个等位基因, 每一个位点标识为AA, BB, CC, AB, BC, AC等等。按Nei等(1979)方法计算遗传距离(Dg)。按Nei(1978)的方法计算观测杂合度(Ho)。杂种优势与亲本遗传距离及子代杂合度的相关分析采用SAS 6.12统计软件进行。
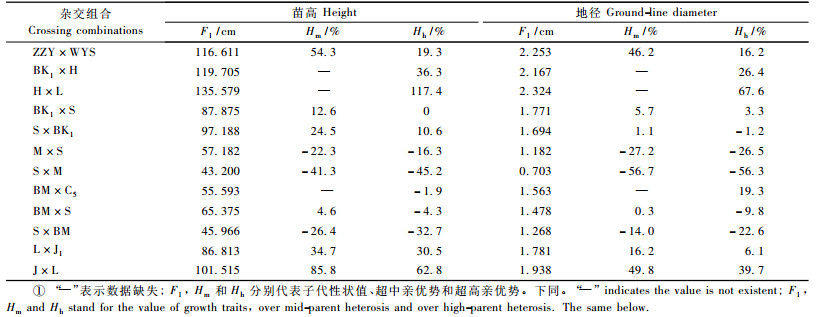
2 结果与分析 2.1 F1杂种优势表现表 1列出了12个杂交组合子代苗期生长量杂种优势表现。苗高的超高亲优势变幅为-45.2%~117.4%, 超中亲优势变幅为-41.3%~85.8%;地径的超高亲优势变幅为-56.3%~67.7%, 超中亲优势变幅为-56.7%~49.8%。由此可以看出, 12个杂交组合子代间苗期生长杂种优势变异较大, 其中有3个杂交组合表现出负向杂种优势, 表明鹅掌楸杂种子代的生长表现取决于亲本选择。
|
|
从176对鹅掌楸属特异性EST-SSR引物中筛选出55对多态性引物对11个交配亲本进行检测(图 1), 55对EST-SSR引物共扩增出60个位点, 每位点等位基因数为2~7个, 共检测到172个等位基因, 每个位点的平均等位基因数为2.9个。

|
图 1 LT018号引物对11个杂交亲本的扩增结果 Figure 1 The amplification products of LT018 locus on 11 parent trees |
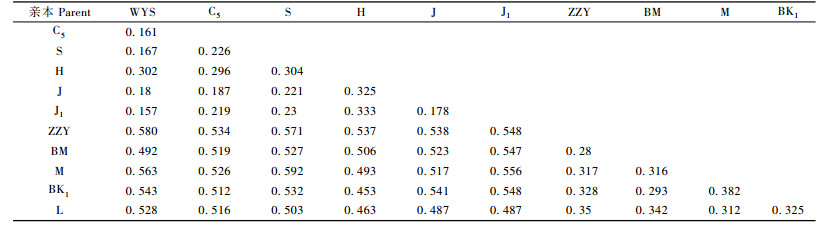
表 2列出了11个交配亲本间的Nei遗传距离。鹅掌楸种内6个亲本间(WYS, C5, S, H, J, J1)的遗传距离变幅为0.157~0.333, 平均为0.232;北美鹅掌楸种内5个亲本间(ZZY, BM, M, BK1, L)遗传距离变幅为0.28~0.382, 平均为0.325;而鹅掌楸与北美鹅掌楸种间亲本的遗传距离变幅为0.453~0.592, 平均为0.529。表明鹅掌楸种间亲本的遗传距离要远远大于种内亲本的遗传距离, 这与鹅掌楸与北美鹅掌楸长期的地理隔离造成的种间遗传分化有关。
|
|
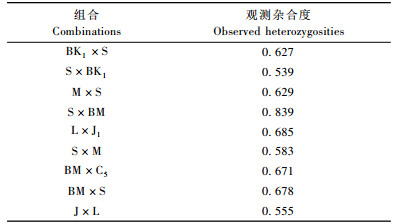
从176对鹅掌楸属特有性EST-SSR引物中筛选出21对多态性引物对9个鹅掌楸杂交组合子代进行检测, 分析其杂合度(表 3)。各杂交组合的观测杂合度变幅为0.539~0.839, 平均为0.645。可以看出各杂交组合杂合度较高, 这与其亲本间较大的遗传距离一致。
|
|
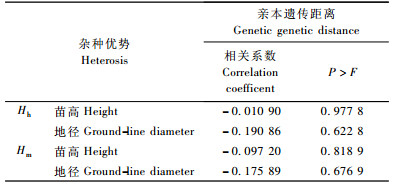
利用SAS6.12统计分析软件将亲本间的遗传距离分别与子代苗高超高亲优势、地径超高亲优势, 苗高超中亲优势和地径超中亲优势进行相关分析(表 4), 结果表明, 杂交子代生长量超高亲优势和超中亲优势与亲本遗传距离间均呈负相关, 但相关性不显著。
|
|
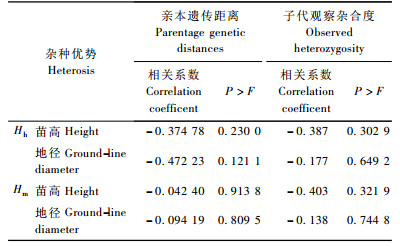
利用SAS统计分析软件将观测杂合度分别与子代苗高超高亲优势、地径超高亲优势, 苗高超中亲优势和地径超中亲优势进行相关分析(表 4), 结果表明, 地径超高亲优势及地径超中亲优势与杂合度之间呈负相关, 但相关性未达显著性水平。
2.5 基于显著影响杂种优势位点的亲本遗传距离与杂种优势相关性分析对每个多态性位点所检测到的观测杂合度与杂种优势进行相关分析(李梅等, 2001), 21个多态性位点中, 显著影响苗高超高亲优势、地径超高亲优势、苗高超中亲优势和地径超中亲优势的位点有5个(α≤0.1)。基于上述5个与杂种优势相关的位点分析亲本遗传距离与杂种优势的相关性(表 5), 结果表明, 亲本遗传距离与杂种优势仍为负相关, 且相关性均不显著。
|
|
对于自交且世代周期短的物种(如农作物), 容易获得纯系亲本, 因而通常以杂种F1与双亲性状进行比较来计算杂种优势。而对于林木, 由于生长周期长、遗传组成高度杂合, 且普遍存在近交衰退, 很难获得纯系亲本, 因此, 采用双亲与杂种F1进行比较来计算杂种优势在林木中很难做到。最佳的方案是对每个杂交亲本采用无性繁殖方法培育亲本无性系苗木, 作为杂交子代杂种优势估算的亲本参照。但这也存在林木成年个体难以进行无性繁殖, 无性系苗木与实生苗木的生理年龄不一致进而影响生长表现等问题。鉴于此, 林木中, 一般以亲本的半同胞子代苗木近似作为亲本对照, 以此来估算杂种优势。
有关鹅掌楸属种间杂种优势的研究报道均以鹅掌楸自由授粉子代为对照(李周岐, 2000), 该方法仅考虑与鹅掌楸亲本的比较而未考虑与北美鹅掌楸亲本的对比。为了更客观地评价杂交鹅掌楸的杂种优势, 本研究利用各杂交组合的亲本自由授粉子代性状的平均值作为参照, 相比以鹅掌楸自由授粉子代为参照, 在一定程度上提高了杂种优势估算的准确性。此外, 由于本试验所涉及的鹅掌楸种间杂交亲本栽植于同一片试验林内, 亲本的自由授粉子代中不可避免地含有一定数量的自然种间杂交子代, 这在一定程度上降低了杂种优势估算值。
3.2 亲本遗传距离与杂种优势相关性关于亲本遗传距离与杂种优势间的关系, 目前尚无定论。有些研究结果表明, 亲本间的遗传距离大则杂种优势强(许占友, 1997; 史延丽等, 2008; Birchler et al., 2003; Stuber et al., 1992; Smith et al., 1990; Lee et al., 1989; Li et al., 2008), 但也有二者不相关的报道(刘宏伟等, 2005; Dudley et al., 1991; Martin et al., 1995; Barbosa et al., 1996; Dhliwayo et al., 2009; Dreisigacker et al., 2005)。在水稻(Oryza sativa)研究中, Zhang等(1995)提出2种异质性, 即一般异质性(general heterozygosity)和特殊异质性(specific heterozygosity), 并发现, 亲本间一般异质性和F1杂种表现的相关性通常较低, 而亲本间的特殊异质性和F1杂种优势呈现极显著正相关。Saghai等(1997)用82个RFLP和26个SSR标记对8个水稻亲本材料进行评价, 发现杂种F1的分子标记杂合度与杂种F1的稻谷产量具有显著正相关。Lee等(1989)在玉米(Zea mays)研究中发现, 亲本间RFLP遗传距离与杂种优势呈显著正相关; Godshalk等(1990)和Dudley等(1991)则认为亲本间RFLP的遗传距离和F1杂种产量的相关性很低。还有研究表明, 分子标记遗传距离和F1表现并非不变的, 而是依赖于被测品系来源。一般地, 亲本来源于相同的杂种优势类群(heterotic group)时亲本遗传距离与F1杂种优势相关性高; 亲本来源于不同的杂种优势类群时, 二者相关性低(Zhang et al., 1995; Boppenmaier et al., 1993; Saghai et al., 1997)。
本文通过对鹅掌楸SSR分子标记亲本遗传距离与杂种子代苗期生长的杂种优势进行的相关分析, 发现二者相关性不显著。其原因可能有2个方面:一方面, 试验所采用的12个F1子代家系均为鹅掌楸与北美鹅掌楸种间杂种, 亲本间的遗传差异较大, 而前人的研究结果表明, 亲本间遗传差异过大时F1杂种优势与亲本间遗传距离相关性低(Boppenmaier et al., 1993; Saghai et al., 1997); 朱其卫(2009)在鹅掌楸种内杂交后代中发现, 其子代的生长表现与亲本遗传距离存在一定程度的相关性(未达显著性水平), 而种内杂交亲本间的遗传距离明显小于种间杂交亲本, 这也为上述推测提供了佐证。另一方面, 试验所采用的SSR分子标记来自于北美鹅掌楸花器官的EST序列, 为有功能基因片段, 但这些片段不一定与控制鹅掌楸生长量的基因有关, 利用SSR标记检测的亲本间遗传距离仅反映亲本间的平均遗传差异状况即亲本间的一般异质性, 而不能确切知道与生长性状有关的具体基因位点的同质或异质性。下一步有必要通过遗传连锁图谱构建和QTL定位, 寻找与生长性状有关或与QTL相连锁的分子标记, 并在此基础上分析亲本遗传距离与杂种优势的相关性。
3.3 子代杂合度与杂种优势的关系目前, 国内外学者对杂种优势形成机制的研究大多集中在亲本遗传距离与子代杂种优势的相关性上, 而对子代杂合度与杂种优势的关系研究较少。原因可能在于前者在实际生产过程中具有更高的实用价值, 可以为亲本选配和杂种优势预测提供依据, 这在生长周期很长的林木上尤为重要。传统的在大量交配组合的基础上通过亲本配合力分析和家系选择及家系内选择进行遗传改良的方法不仅费时而且费力, 从亲本遗传距离和杂种优势的关系出发进行交配设计, 这对于缩小杂交试验规模、提高杂交育种效率和加快良种化进程具有重要的意义。
子代的遗传多样性来源于亲本间的遗传差异, 遗传差异不仅包括父母本之间的遗传距离也包括父母本各自的遗传组成。一般来说, 分布地区距离较远、类型不同的种群间杂交, 以及长期与外界隔绝、在遗传组成上基因杂合度较高的种群间杂交, 子代可以获得较大的遗传多样性(王明庥, 2001)。一般地, 亲本间的遗传距离与子代遗传多样性相关性较强。但应注意的是, 不论是通过形态标记、生化标记还是分子标记来估算亲本间的遗传距离, 都只能反映亲本间的平均遗传差异状况, 而不能确切知道某个基因位点的同质或异质性(李周岐等, 2002)。因此, 亲本间遗传差异并不能直接反映出子代基因组的杂合性, 因为在亲、子基因传递过程中还存在基因分离与重组, 因而, 利用亲本遗传距离来衡量子代杂合性, 这在一定程度上势必会出现偏差。
为客观地分析鹅掌楸子代杂合性与杂种优势之间的关系, 本文利用SSR分子标记对其作了进一步探讨。从研究结果来看, 杂种优势与子代杂合度相关性也不显著, 表明亲本遗传距离和杂交子代杂合度可能并非鹅掌楸杂种优势形成的主要原因。当然, 本文研究的是杂交鹅掌楸苗期生长的杂种优势表现, 成年期生长性状的杂种优势表现可能会发生波动, 从而改变杂种优势与亲本遗传距离/子代杂合度之间相关系数的大小及显著性水平, 对此, 将进行跟踪研究。
兰进好, 张宝石, 周鸿飞. 2005. 作物杂种优势遗传基础研究进展[J]. 中国农学通报, 21(1): 114-119. |
李梅, 施季森, 甘四明, 等. 2001. 杉木杂交亲本分子遗传变异与子代生长相关的研究[J]. 林业科学研究, 14(1): 35-40. |
李周岐. 2000. 鹅掌楸属种间杂种优势的研究. 南京林业大学博士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-HBLY401.017.htm
|
李周岐, 王章荣. 2002. 用RAPD标记进行鹅掌楸杂种识别和亲本选配[J]. 林业科学, 38(5): 169-174. DOI:10.11707/j.1001-7488.20020529 |
刘宏伟, 刘秉华, 张改生, 等. 2005. RAPD分子标记与小麦杂种优势相关性研究[J]. 麦类作物学报, 25(6): 1-5. DOI:10.7606/j.issn.1009-1041.2005.06.231 |
卢庆善, 孙毅, 华泽田. 2002. 农作物杂种优势[M]. 北京: 中国农业科技出版社: 11-20.
|
史延丽, 王坚, 李自超. 2008. 粳稻分子标记遗传分化与杂种优势关系的研究[J]. 西北农业学报, 17(5): 166-170. |
王明庥. 2001. 林木遗传育种学[M]. 北京: 中国农业出版社: 57-64.
|
王章荣. 1997. 中国马褂木遗传资源的保存与杂交育种前景[J]. 林业科技通讯, (9): 8-10. |
许占友. 1997. 小麦分子遗传差异与杂种优势关系及育性恢复基因分子标记的研究. 中国农业大学博士学位论文.
|
叶金山, 王章荣. 2002. 杂种马褂木杂种优势的遗传分析[J]. 林业科学, 38(4): 67-71. DOI:10.11707/j.1001-7488.20020411 |
张博, 张露, 诸葛强, 等. 2004. 一种高效的树木总DNA提取方法[J]. 南京林业大学学报:自然科学版, 28(1): 13-17. |
张晓飞. 2008. 鹅掌楸不同交配组合子代幼龄期生长性状的遗传分析. 南京林业大学硕士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-LHYS503.032.htm
|
朱其卫. 2009. 鹅掌楸不同交配系统子代遗传多样性研究. 南京林业大学硕士学位论文. http://life.scichina.com:8082/sciC/CN/abstract/abstract520135.shtml
|
Barbosa Neto J F, Sorrells M E, Cisar G. 1996. Prediction of heterosis in wheat using coefficient of parentage and RFLP-based estimates of genetic relationship[J]. Genome, 39: 1142-1149. DOI:10.1139/g96-144 |
Birchler J A, Auger D L, Riddle N C. 2003. In search of a molecular basis of heterosis[J]. The Plant Cell, 15(10): 2236-2239. DOI:10.1105/tpc.151030 |
Boppenmaier J, Melchinger A E, Seiltz G. 1993. Genetic diversity for RFLPs in European maize inbreds Ⅲ.Performance of crosses within versus between heterotic groups for grain traits[J]. Plant Breeding, 111(3): 217-226. DOI:10.1111/pbr.1993.111.issue-3 |
Dhliwayo T, Pixley K, Menkir A, et al. 2009. Combining ability, genetic distances, and heterosis among elite CIMMYT and IITA tropical maize inbred lines[J]. Crop Sci, 49(4): 1201-1210. DOI:10.2135/cropsci2008.06.0354 |
Dreisigacker S, Melchinger A E, Zhang P, et al. 2005. Hybrid performance and heterosis in spring bread wheat, and their relations to SSR-based genetic distance and coefficients of parentage[J]. Euphytica, 144(1/2): 31-38. |
Dudley J W, Saghai Maroof M A, Rufener G K. 1991. Molecular marker and grouping of parents in maize breeding programs[J]. Crop Sci, 31(3): 718-723. DOI:10.2135/cropsci1991.0011183X003100030036x |
Godshalk E B, Lee M, Lamkey K R. 1990. Relationship of restriction fragment length polymorphisms to single-cross hybrid performance of maize[J]. Theor Appl Genet, 80(2): 273-280. |
Lee M, Godshalk E B, Lamkey K R, et al. 1989. Association of restriction fragment length polymorphisms among maize inbreds with agronomic performance of their crosses[J]. Crop Sci, 29(4): 1067-1071. DOI:10.2135/cropsci1989.0011183X002900040050x |
Li Xiaojie, Yang Guanpin, Shi Yuanyuan, et al. 2008. Prediction of the heterosis of Laminaria hybrids with the genetic distance between their parental gametophyte clones[J]. Journal of Applied Phycology, 20(6): 1097-1102. DOI:10.1007/s10811-008-9321-9 |
Martin J M, Tablber L E, Lanning S P. 1995. Hybrid performance in wheat as related to parent diversity[J]. Crop Sci, 35(1): 104-108. DOI:10.2135/cropsci1995.0011183X003500010019x |
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals[J]. Genetics, 89(3): 583-590. |
Nei M, Li W H. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases[J]. Proc Natl Acad Sci, 76(10): 5269-5273. DOI:10.1073/pnas.76.10.5269 |
Saghai Maroof M A, Yang G P, Zhang Q F, et al. 1997. Correlation between molecular marker distance and hybrid performance in U[J]. S.southern long grain rice.Crop Sci, 37(1): 145-150. |
Smith O S, Smith J S C, Bowen S L, et al. 1990. Similarities among a group of elite maize inbreds as measured by pedigree, F1 grain yield, grain yield heterosis, and RFLPs[J]. Theor Appl Genet, 80(6): 833-840. |
Stuber C W, Lincoln S E, Wolff D W, et al. 1992. Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers[J]. Genetics, 132: 823-839. |
Xu Meng, Li Huogen, Zhang Bo. 2006. Fifteen polymorphic simple sequence repeat markers from expressed sequence tags of Liriodendron tulipifera[J]. Molechlar Ecology Notes, 6(3): 728-730. DOI:10.1111/men.2006.6.issue-3 |
Zhang Q F, Gao Y J, Saghai Maroof M A. 1995. Molecular divergence and hybrid performance in rice[J]. Molecular Breeding, 1(2): 133-142. DOI:10.1007/BF01249698 |