 2011, Vol. 47
2011, Vol. 47文章信息
- 王秀花, 马雪红, 金国庆, 陈柳英, 周志春
- Wang Xiuhua, Ma Xuehong, Jin Guoqing, Chen Liuying, Zhou Zhichun
- 木荷天然林分个体类型及材性性状变异
- Variation Pattern of Individual Types and Wood Characters in Natural Stands of Schima superba
- 林业科学, 2011, 47(3): 133-139.
- Scientia Silvae Sinicae, 2011, 47(3): 133-139.
-
文章历史
- 收稿日期:2009-05-31
- 修回日期:2009-10-13
-
作者相关文章



2. 福建省建瓯市林业科技推广中心 建瓯 353100
2. Extending Center for Forestry S & T of Jian'ou, Fujian Province Jian'ou 353100
较之于作物,林木存在更为丰富的自然变异,表现在生长、形态、生理、材性等方面。人们所观测到表型变异其实是基因型与环境互作的结果,可在一定程度上反映遗传变异的大小和规律,尤其是对于缺乏遗传背景研究的一些树种,通过种内表型变异的研究,可以较好地说明其遗传改良的潜力。李文英等(2005)研究发现蒙古栎(Quercus mongolica)天然表型丰富,随着经纬度的增加,坚果逐渐增大,坚果形状由近球形向长椭球形变异,叶形由长倒卵形向宽倒卵形变异。周志春等(1995)分析测试了马尾松(Pinus massoniana)自然分布区内14个天然群体的木材化学组分和浆纸性能,发现其木材戊糖、灰分含量,纸张抗张、耐破和撕裂强度呈现随纬度明显变化的地理模式,与经度无关。Scheiner (1993)认为在各种不同环境中等位基因表达的差异及位点间相互作用的变化都有可能引起物种表型的变异。结合所处生境的差异,对林木形态分化、变异和生长特性进行研究有助于了解林木沿生境梯度的变异幅度和规律,从而进行有效的遗传改良。
木荷(Schima superba)是山茶科(Theaceae)常绿大乔木,具有生长迅速、材质优异、少病虫害、对立地要求不严等优良特性,不仅是我国南方各省区生物防火林带建设的主要树种和高效的生态树种,还是造林成效好、速生珍优的阔叶用材树种(阮传成等,1995)。木荷自然分布广泛,种内变异极其丰富(张萍等,2006; 金则新等,2007),研究发现来自不同产地的种源存在显著的遗传差异并多呈纬向变异模式(张萍等,2004; 周志春等,2006)。此外木荷天然林分内的变异也非常丰富,存在众多的个体变异类型,如木材颜色有红白之分,树皮厚度有厚薄、树皮颜色有黑灰、树皮形状有光滑、鳞状、条状之分等,不同类型的个体其生长特性和材质材性明显不同。研究组从2001年开始了以优质用材和生态高抗为目标的木荷育种技术研究,取得了一些阶段性的研究成果,但有关木荷个体类型和材性变异的研究尚未涉及。本文在福建和浙江2省选择不同纬度和海拔梯度的木荷天然林分,系统开展木荷个体类型调查和材性分析,以揭示树皮厚薄/形状/颜色、木材纹理扭曲度、木材颜色等个体类型和木材基本密度在天然林分内的表型变异规律及产地纬度与海拔的影响,从而为木荷种内变异的发掘和利用提供科学理论依据。
1 材料与方法 1.1 天然林分选择为研究木荷天然林分个体类型和材性表型变异规律及产地纬度与海拔的影响,在浙江和福建2省的木荷主要分布区,分别按低海拔(200 ~ 400 m)和高海拔(600 ~ 800 m)选择2组不同纬度梯度(经度相似)的木荷天然林分,即低海拔组包括福建建瓯徐墩(27°15′N,118°11′E)、浙江龙泉(28°04′N,119°06′E)和淳安(29°28′N,119°08′E)3个天然林分,高海拔组包括福建尤溪(26°07′N,118°20′E)、建瓯顺阳(27°01′N,118°37′E)和浙江庆元(27°40′N,119°00′E) 3个天然林分,具体见表 1。在上述选择的6个天然林分中,福建建瓯的2个林分及浙江龙泉与庆元的2个林分又可构成不同海拔梯度(经纬度相同)的2组。选择的林分要求为当地起源的以木荷为主的天然林分,面积要求在6 hm2以上,年龄在30年以上。
|
|
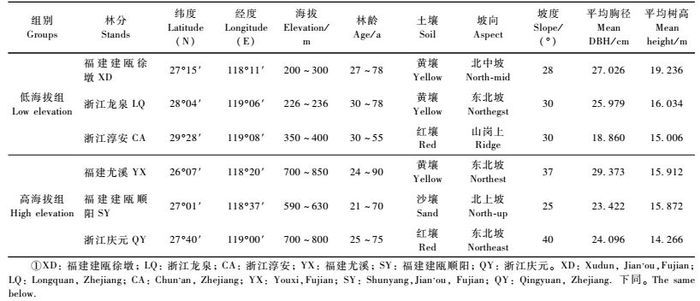
在每一木荷天然林分的代表性地段设置一个0.1 hm2大小的临时样地,随机选择50株木荷优势木和亚优势木,全面量测其树高、胸径、通直度(按通直、较通直、一般、弯曲、严重弯曲5级,分别记分为5,4,3,2和1,分数越高越通直)、圆满度(用树干下部2 m高处直径与胸径之比表示)、树皮和木材颜色(分别为灰绿、灰棕、棕色和浅棕、棕红、红褐色,分别用1,2,3表示,且由1到3表示颜色由浅到深,取木芯时观察,采用欧洲皇家比色系卡比色)、树皮形状(光滑、鳞片、长条状分别用1,2,3表示)、树皮厚度、木材纹理扭曲度(以木材纹理偏离树干主轴角度的正切值表示)等,并在每株样木胸高处上坡方位用6 mm直径的生长锥钻取一根髓心至树皮的完整无疵木芯。对所取的木芯自髓心向外,前40轮按每5个年轮切成一段,共8个年轮段(R1 ~R8),40轮以后归为一段(RX),分别量测各年轮段的年轮宽度(Wi),并用最大含水量法(Smith,1954)测定每年轮段的木材基本密度(Di)。
1.3 数据分析利用各年轮段宽度(Wi)测定值估算其所代表的年轮段面积(Ai),进而计算每根木芯前30轮的加权平均基本密度BD30 = Σ AiDi /Σ AiWi(i = 6)和平均年轮宽度BW30 = Σ Wi /30(i = 6),以消除树龄差异的影响。采用SAS8.01软件进行方差分析和相关分析,并采用Excel软件计算变异系数。数据分析时,树干通直度、树皮颜色、木材颜色、树皮形状的数值经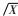
对木荷天然林分各表型性状进行方差分析(表 2),结果表明除树干纹理扭曲度外,木荷树干通直度和圆满度、树皮厚薄/形状/颜色、木材颜色、木材基本密度等性状在不同纬度和海拔的天然林分间均存在显著的表型差异,如林分间树皮厚度变幅为0.753 ~ 1.070 mm,最大值较最小值高出42.1%,1 ~ 30轮木材基本密度和平均年轮宽度在林分的变幅分别为0.523 ~ 0.589 g·cm-3和2.083 ~ 2.760mm,最大值分别大于最小值12.6%和32.5%,这说明木荷天然林分间存在丰富的表型变异。
|
|
进一步分析(表 2)发现,木荷树干形质和木材基本密度等性状在天然林分内个体间也存在丰富的变异,其中树干纹理扭曲度、树皮厚薄/形状/颜色、木材颜色和1 ~ 30轮平均年轮宽度在林分内个体间的变异较大,如树干纹理扭曲度个体变异系数平均值高达70.67%,这一结果意味着对这些性状进行个体选择或优树选择的必要性及巨大的选择潜力; 相对而言,木荷树干圆满度、通直度和1 ~ 30轮木材基本密度在林分内个体间变异较小,如树干圆满度和1 ~ 30轮木材基本密度的平均个体变异系数仅分别为5.43%和7.15%,说明木荷天然林分内优势木或亚优势木皆具有树干通直、圆满的优良特点,个体间木材密度差异较小。通过比较可看出,各林分内的表型多样性差异较大,总体来讲以福建尤溪、建瓯顺阳2林分内的表型变异最为丰富,2林分表型性状变异系数依性状不同,分别变化在5.64% ~73.97%和6.32% ~ 76.93%间,而浙江淳安林分内的变异最小,表型性状变异系数依性状不同变化在4.58% ~ 64.49%间。
木荷个体存在很多类型,尤其是其树皮形状、树皮颜色、木材颜色等类型丰富。现将木荷天然林个体树皮颜色归为灰绿、灰棕和棕黑色3类,树皮形状有光滑、鳞片、条状3种,木材颜色划为浅棕色、棕红色和红褐色3类,以分别计算出6个天然林分中各类型个体所占比例。
从表 3看出:树皮颜色在各林分内以棕黑色为主,在浙江淳安、福建尤溪和建瓯顺阳3个林分中比例都在60%以上; 树皮形状除浙江淳安、福建尤溪2林分以条状为主外,其他林分均以鳞片状为主,如在福建建瓯林分中过半树木的树皮为鳞片状,树皮光滑的个体类型比例较少。从浙江淳安和福建尤溪林分个体树皮的颜色、形状所占比例来看,树皮颜色和形状有很大的相关性,越粗糙的树皮其颜色越深。木材颜色在各林分内浅棕色类型所占比例最大,在中、高纬度地区的浙江龙泉、浙江淳安和浙江庆元3个林分内所占比例达90%以上,然而在低纬度地区的天然林分中,木材颜色变深,棕红色和红褐色木材类型出现的比例较大,特别是在低纬度的福建尤溪林分中,红褐色木材类型的比例达27.5%。
|
|

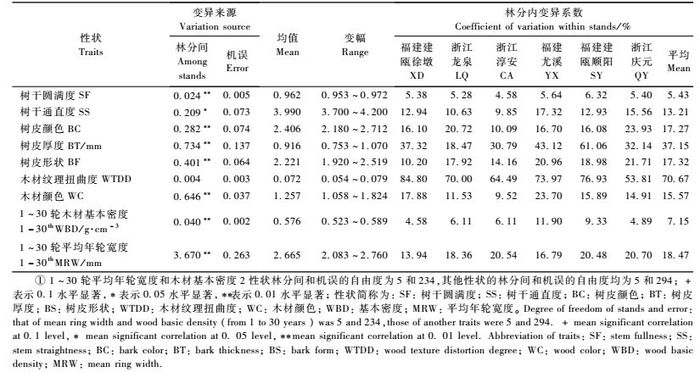
由图 1A所示:木荷天然林木材基本密度径向变异模式符合Panshin等(1980)所述的第Ⅲ种类型,即由髓心向树皮方向逐渐减小。其中低海拔组的福建建瓯徐墩、浙江龙泉、淳安3个天然林分木材基本密度下降趋势较缓,5 ~ 10轮间虽然下降剧烈,但10轮以后变化趋于平缓,而高海拔组的福建尤溪、建瓯顺阳、浙江庆元3个天然林分其木材基本密度自髓心至树皮以直线形式急剧降低,这说明高海拔林分的木材基本密度径向变异较低海拔林分强烈。由图 1A还可以看出:木荷髓心处的木材基本密度远高于边材部分,可能与心材浸提物的沉积有关(徐有明等,2002; 李莉等,2008)。福建尤溪天然林分的木材基本密度明显大于其他地点的林分,这与林龄大有关。

|
图 1 木荷天然林分木材基本密度、年轮宽度的径向变异 Figure 1 The radial variation of wood basic density and ring width in natural stands of S.superba |
木荷天然林分年轮宽度的径向变异不同于木材基本密度(图 1B),除浙江淳安林分外,一般由髓心向树皮方向年轮先变宽后变窄。木荷幼林直径生长缓慢,在髓心附近的年轮宽度较窄,5 ~ 15轮间直径生长加快,15 ~ 25轮间为平稳生长期,年轮最宽,25轮之后年轮宽度逐渐下降。在高海拔组,福建尤溪林分直径的快速生长期(5 ~ 20轮)较之于其他2林分(10 ~ 15轮)要长,平稳生长期达10年(20 ~ 30轮),这与其所处纬度较低有关; 与高海拔组林分比较,低海拔组3个天然林分的直径平稳生长期较长,为15年(15 ~ 30轮)左右,说明低海拔较高海拔林分直径生长快,平稳生长期长。
2.3 海拔、纬度对木荷天然林树干形质和木材基本密度等影响表 4按产地纬度和海拔分组列出了木荷天然林分树干形质和木材基本密度等测定值。结果表明:木荷天然林分的树干形质和木材基本密度等与所处产地的纬度和海拔有关。产地纬度对各性状的影响存在较大差异,不管是低海拔组还是高海拔组的林分,来自较高纬度地区的天然林分呈现树干圆满度和通直度增大、树皮厚度变薄、木材颜色变淡的趋势。然而产地纬度对木荷天然林树皮颜色、树皮形状、木材基本密度和年轮宽度的影响却较为复杂,未发现明显的规律性; 相对于产地纬度,海拔影响的规律性却较为明显,来自较高海拔的木荷天然林分具有树干圆满,树皮薄而光滑,树干通直度低、年轮窄、木材基本密度小等趋势,但树皮和木材颜色在不同海拔间差异较小。
|
|
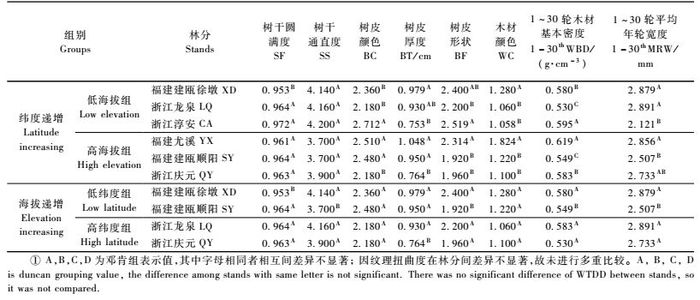
选取木荷中心产区福建建瓯的低海拔(徐墩)和高海拔(顺阳) 2个代表性天然林分,分别对其林分内个体形质和木材基本密度等进行相关分析。表 5结果表明:木荷天然林分个体性状间的相关性因林分海拔不同而有较大差异。对于处于较高海拔的建瓯顺阳木荷天然林分,个体树皮的颜色、厚度和形状间关系密切,其间的相关系数都达1%显著性水平,树皮颜色深的个体其树皮较厚且多为条状。根据树皮厚度和颜色还能较好地预测其木材色泽、密度大小和径生长情况(其间相关系数都达1%或5%显著性水平),树皮厚、颜色深的个体其木材颜色深、木材基本密度大、径生长快,可见对于较高海拔的木荷天然林分,树皮性状是较好指示材性和径生长的形态指标,对木荷优树选择可提供较好的帮助。此外还发现,个体树干圆满度和通直度与树皮的颜色和形状、木材的基本密度和年轮宽度相关系数均达到5%显著性水平,树干圆满的个体树皮相对光滑、颜色较浅、木材基本密度较小,而树干越通直,其树皮颜色则越深、径生长也越快。树干纹理扭曲度与其他性状互为独立,其间的相关性较小。
|
|

对于地处较低海拔的建瓯徐墩木荷天然林分,性状相关分析发现树皮厚度是一个较好指示树皮其他性状和木材基本密度的指标(相关系数达到10%或5%显著性水平),树皮较厚的个体其树皮颜色和木材颜色都较深,木材基本密度较大,但与高海拔林分不同的是其树皮较为光滑。木材基本密度分别与树皮厚度和形状呈5%显著性水平的正相关和负相关,树皮厚且光滑的个体其木材基本密度相对较高。此外,相关分析还发现,年轮宽度也即个体的径生长与树干圆满度和通直度、树皮颜色、纹理扭曲度等呈5%显著性相关,林分内径生长量大的个体其树干通直、树皮颜色较浅、纹理扭曲性较小,但其树干圆满度却较低,这一结果对于在较低海拔木荷天然林内开展优树选择具有很好的指导意义,选择树干通直、树皮颜色浅、径生长量大的木荷优树,其木材往往有纹理扭曲度和基本密度变小的趋势。
3 结论与讨论 3.1 木荷天然林分个体类型和材性的表型变异本文在浙江和福建2省选择成一定纬度和海拔梯度的6个木荷天然林分进行个体类型调查和材性测试,发现除木材纹理扭曲度外,树干形质、树皮形态和木材基本密度等性状在林分间存在显著的表型变异,而这种表型变异主要受林分所处纬度和海拔影响,说明木荷种源显著的纬向变异有其必然的基础(张萍等,2004; 周志春等,2006)。试验发现来自较高纬度的木荷林分其树干较为通直圆满、树皮较薄、木材颜色较浅,其他性状随产地纬度的变化规律不明显。较之于产地纬度,海拔影响更具规律性,随着海拔升高,木荷天然林分具有树干圆满,树皮薄而光滑,但树干通直度低、年轮窄、木材基本密度小等变化趋势。木荷天然林不仅在林分间存在显著的表型变异,而且在林分内个体变异更为丰富,其中树皮厚度、颜色、形状及木材颜色等类型多样,木材纹理扭曲度、树皮厚薄/形状/颜色、木材颜色和年轮宽度的个体变异系数在15% ~ 70%间。然而因木荷具有树干通直圆满、个体间木材密度差异较小的优良特点,其林分内个体间变异系数较小,这些特性有利于速生、优质(树干通直圆满、纹理直、树皮薄、材色深)工艺用材品种的定向选育。木荷个体类型尤其是树皮形态/颜色还与树龄有关,根据大量的观测资料发现木荷在30年生左右其个体类型就已经稳定。本文所选的6个木荷天然林分林龄基本一致,多数个体的年龄在30 ~ 40年间,这意味着林龄对个体类型变异影响可以忽略,个体类型变异主要与林分所处纬度和海拔有关,如试验观测到地处中高纬度的林分木材颜色浅,浅棕色木材的个体达到90%以上,地处低纬度的林分木材颜色较深,棕红色和红褐色木材的个体类型出现的比例较大,这对材色的品种选育有很好的指导意义。
3.2 木荷天然林分生长、形质和木材基本密度的表型相关木荷个体类型多样,树干形质、树皮形态和木材密度等性状间存在不同程度的相关性且与林分所处产地环境有关。本文选用木荷中心产区福建建瓯2个不同海拔的天然林分进行个体性状相关分析,结果发现:在较高海拔的林分内个体树皮性状与材性和径生长相关密切,树皮厚、颜色深的个体其木材颜色也深、木材基本密度大、径生长快,因此在较高海拔的木荷天然林分内可根据个体树皮的厚薄和颜色对材性和径生长进行间接选择; 在较低海拔的林分内,树皮厚度对木材性质也有较好的指示作用,薄皮型个体的树皮和木材颜色浅、木材密度大,而厚皮类型个体的树皮和木材颜色深、木材密度小,此外相关分析还发现树干通直、树皮颜色浅、径生长量大的个体木材纹理直,且木材密度有增加趋势。因此,在进行木荷优树选择时可根据树皮厚度、颜色等对材性进行间接选择。
3.3 木荷天然林分年轮宽度和木材基本基本密度的径向变异根据材性差异可将木材分成幼龄材和成熟材2部分,近髓心一定年轮内的木材为幼龄材(李坚,2001)。与成熟材比较,幼龄材的纤维短,细胞壁薄、壁腔大、木材密度小,多数松类和软阔叶树种的木材密度从髓心至树皮逐渐增大,到达一定年龄后渐趋稳定或略有降低(Zobel et al., 1989)。木荷是广泛分布于我国亚热带地区的珍贵硬阔叶类树种,其木材基本密度的径向变异不同于松类和软阔叶类树种,近髓心部分的木材密度最大,由髓心向树皮不断减小,类似于云杉(Picea asperata) (罗建勋等,2004)、日本花柏(Chamaecyparis pisifera) (王大鹏,2007; 徐有明等,2006)及西班牙栎(Quercus falcata)等硬阔叶类树种(Hamilton,1961),这与其近髓心部也即心材部分有大量的浸提物沉积有关。研究还发现木荷不同天然林分木材基本密度径向变异也存在较大的差异,如福建建瓯顺阳林分的木材基本密度从髓心向树皮急剧减小,而如浙江淳安等林分的木材基本密度由髓心向树种变化较缓,径向均一性较高,这为选择材性径向均一性高的种源和个体提供了可能。木荷天然林分的年轮宽度则遵循多数树种的径向变异规律(姜笑梅等,2003; 费本华等,2000; 2005),即由髓心向树皮年轮先变宽后变窄,在15 ~25轮平稳生长期时年轮最宽,进入成熟期后年轮又渐变窄。同样,不同林分间仍存在一定差异,发现来自较低纬度和较低海拔的林分年轮较宽,平稳生长期较长。
费本华, 高慧, 丁佐龙. 2000. 银杏木材的密度和化学性质[J]. 东北林业大学学报, 28(4): 47-49. |
费本华, 江泽慧, 虞华强, 等. 2005. 人工经济林木材性质研究[J]. 林业科学, 41(1): 116-122. DOI:10.11707/j.1001-7488.20050120 |
姜笑梅, 殷亚方, 浦上弘幸. 2003. 北京地区Ⅰ-214杨树木材解剖特性与基本密度的株内变异及其预测模型[J]. 林业科学, 39(6): 116-121. |
金则新, 李钧敏, 蔡琰琳. 2007. 不同海拔高度木荷种群遗传多样性的ISSR分析[J]. 生态学杂志, 26(8): 1143-1147. |
李坚. 2001. 木材科学[M]. 北京: 高等教育出版社.
|
罗建勋, 李晓清, 孙鹏, 等. 2004. 云杉天然群体管胞和木材基本密度性状变异的研究[J]. 北京林业大学学报, 26(6): 80-85. |
李文英, 顾万春. 2005. 蒙古栎天然群体表型多样性研究[J]. 林业科学, 41(11): 49-56. |
李莉, 王昌命. 2008. 工业林木荷木材化学成分及其变异的研究[J]. 山东林业科技, 38(2): 5-8. |
王大鹏. 2007. 日本花柏木材管胞解剖特征与基本密度的变异. 华中农业大学农学硕士学位论文, 40-43. http://cdmd.cnki.com.cn/Article/CDMD-10504-2007209838.htm
|
徐有明, 林汉, 江泽慧, 等. 2002. 橡胶树生长轮宽度、木材密度变异及其预测模型的研究[J]. 林业科学, 38(1): 95-102. DOI:10.11707/j.1001-7488.20020115 |
徐有明, 史玉虎, 王大鹏, 等. 2006. 日本花柏人工林生长规律与晚材率、木材密度的变异[J]. 东北林业大学学报, 34(1): 48-51. |
阮传成, 李振问, 陈诚和, 等. 1995. 木荷生物工程防火机理及应用[M]. 成都: 电子科技大学出版社.
|
张萍, 金国庆, 周志春, 等. 2004. 木荷苗木性状的种源变异和地理模式[J]. 林业科学研究, 17(2): 192-198. |
张萍, 周志春, 金国庆, 等. 2006. 木荷种源苗高生长参数变异研究[J]. 林业科学研究, 19(1): 61-65. |
周志春, 范辉华, 金国庆, 等. 2006. 木荷地理遗传变异和优良种源初选[J]. 林业科学研究, 19(6): 718-724. |
周志春, 秦国峰, 李光荣, 等. 1995. 马尾松天然林木材化学组分和浆纸性能的地理模式[J]. 林业科学研究, 8(1): 1-6. |
Hamilton J K. 1961. Variation of wood properties in southern red oak[J]. For Prod J, 11: 267-271. |
Panshin A J, Card de Zeeuw. 1980. Textbook of wood technology. 4th edition. New York: McGraw Hillbook Company.
|
Scheiner S M. 1993. Genetics and evolution of phenotypic plasticity[J]. Annual Review of Ecology and Systematics, 24(1): 35-68. DOI:10.1146/annurev.es.24.110193.000343 |
Smith D M. 1954. Maximum moisture content method for determining specific gravity of small wood samples. USDA For Serv For Prod Lab Rep, 2014.
|
Zobel B J, Buijtenen J P. 1989. Wood variation Its causes and control. Berlin: Springer Verlage, 393.
|