 2011, Vol. 47
2011, Vol. 47文章信息
- 何善勇, 朱银飞, 阿地力·沙塔尔, 温俊宝, 陈梦, 田呈明
- He Shanyong, Zhu Yinfei, Adili Satar, Wen Junbao, Chen Meng, Tian Chengming
- 枣实蝇在中国的风险评估
- Pest Risk Assessment of Carpomya veusuviana in China
- 林业科学, 2011, 47(3): 107-116.
- Scientia Silvae Sinicae, 2011, 47(3): 107-116.
-
文章历史
- 收稿日期:2009-09-24
- 修回日期:2010-06-05
-
作者相关文章



2. 新疆农业大学 乌鲁木齐 830052;
3. 新疆林业有害生物防治检疫总站 乌鲁木齐 838000
2. Xinjiang Agriculture University Urumqi 830052;
3. Xinjiang Forest Diseases and Insect Pests Control and Quarantine Station Urumqi 830052
枣实蝇(Carpomya vesuviana)属双翅目(Diptera)实蝇科(Tephritidae)实蝇亚科(Trypetina)实蝇族(Trypetini)咔实蝇属(Carpomya),2007年5月被列入我国进境植物检疫性有害生物名录,2008年增列为全国林业检疫性有害生物,为枣属(Zizyphus)植物重大害虫。在源发地印度,枣实蝇是枣树最具毁灭性的害虫(Lakra et al., 1983),几乎所有种植枣树的地区均有枣实蝇危害,局部地区危害率高达100% (Pareek et al., 2003),已成为印度枣产业发展的主要限制因子(Lakra et al., 1983)。我国于2007年在新疆吐鲁番地区初次发现该虫,当年全地区1 /3的结果枣树受害,损失十分惨重(阿地力·沙塔尔等,2008)。
目前枣实蝇仅传入我国新疆吐鲁番地区,除新疆外,我国还有众多枣产区,故枣实蝇对我国枣产业构成巨大潜在威胁,开展其潜在适生区预测意义重大。吕文刚等(2008)利用CLIMEX2.0软件对枣实蝇在我国的潜在适生区进行了预测,但预测所用的枣实蝇发育生物学数据均来自于国外资料。本文通过在吐鲁番地区的实地观察试验,获取了第一手枣实蝇生物生态学特性资料,同时结合CLIMEX3.0和GARP软件对枣实蝇在我国的潜在适生区进行预测。根据预测结果,利用有害生物危险性量化评估指标,对枣实蝇在各省市区的潜在危险性进行评估,以期为相关部门提供决策依据。
1 材料与方法 1.1 寄主分布资料枣树分布资料主要来源于中国种植信息网(http://zzys.agr.igov.cn),统计近5年全国枣种植产量确定枣产区最新种植面积(周国梁等,2007)。根据省区的行政面积推算出枣果产量密度,藉此确定枣树在我国各分布区内种植密度:0 kg·km-2<枣果产量<500 kg·km-2,则认为该地区为低密度种植区; 500 kg·km-2≤枣果产量<1 000 kg·km-2,则为中等密度种植区; 枣果产量≥1 000 kg·km-2,为高密度种植区。对于该网无记录的地区,则通过查阅相关文献和其他网上资源获取。
1.2 利用CLIMEX3.0进行适生性分析 1.2.1 软件介绍本研究采用澳大利亚Hearne Scientific Software公司出版的CLIMEX3.0软件。同2.0版本相比,3.0版本在地图容量、地图构建、地图打印、结果表格及图形输出、气象数据输入等方面做了相应改善(如地图采用了标准的shape文件格式,输出结果表格和图形可导出为栅格文件格式,便于与GIS软件结合进行结果图形的处理)。主要模块仍为地区模拟和气候相似性比较。本研究主要采用模拟功能中的单物种地区比较。
1.2.2 基础资料收集与整理1) 枣实蝇基础生物学吐鲁番地区枣实蝇越冬蛹的发育起点温度C = (13.63 ± 1.91) ℃,有效积温K = (807.55 ± 110.12)日度,在33 ℃条件下发育历期最短(何善勇等,2009a)。在吐鲁番地区,枣实蝇越冬蛹生存的土壤平均温度最低为-8 ℃ (何善勇等,2009b)。Sangwan等(1992)在印度研究发现40 ℃时枣实蝇蛹50天没有羽化现象。Dashad等(1999)在印度Bawal地区研究发现最高温度17 ~ 25 ℃、最低温度2.3 ~ 4.8 ℃、最适相对湿度62% ~ 85.5%时枣实蝇危害率较高。Lakra等(1985)在印度Hisar地区发现温度低于5 ℃时,各虫态发育历期均会推迟,温度高于40 ℃,特别是45 ℃时,伴随着20% ~ 30%的空气相对湿度,对成虫有致死作用,周平均最低温度10 ~ 25 ℃、最高温度25 ~ 40 ℃、相对湿度25% ~75%被认为是最适合枣实蝇生存的气候条件,周平均20 ~ 40 mm的中等降雨量有助于枣实蝇种群数量的增加,50 ~ 120 mm中到大雨会影响枣实蝇羽化率,甚至引起成虫死亡,降雨量和相对湿度在决定枣实蝇发生率上起着重要作用。
2) 中国气象数据收集根据国家气象信息中心提供的中国地面气候标准值月值数据集(1971—2000),结合CLIMEX自带的85个气象站点,整理得到全国740个气象站点的月均最高气温、月均最低气温、月均相对湿度和月累积降雨量等关键气候数据。
1.2.3 分析方法1) 最优参数值设置已知的枣实蝇生物生态学资料有限,尤其是环境因子交互作用影响枣实蝇发生的资料极少,因此分析时未设置环境交互作用参数项,而选择了主要的17个参数项。参数设置时,先根据试验和整理得到的枣实蝇基础生物学数据初步确定各参数值。运行软件并反复调试各参数值,对枣实蝇在全球的分布进行模拟,直到模拟结果与枣实蝇实际情况根据(尤其是源发地印度的分布点),达到最大程度的拟合,由此确定预测最终所用参数值。
2) 生态气候EI值划分生态气候指数EI的取值范围为0 ~ 100,其大小代表物种在目标区域内的潜在适生程度。CLIMEX认为:EI值为0则该地不适合该物种生存; EI值大于30时,表示该地区非常适合物种生存(Sutherst et al., 1999)。根据CLIMEX预测我国枣实蝇已知发生地吐鲁番市、鄯善县和托克逊县的EI值,结合枣实蝇在当地的实际发生情况(阿地力·沙塔尔等,2008),将预测结果EI值划分为4个水平:“EI = 0”表示该地不适生; “0<EI≤10”表示该地低度适生; “10<EI<30”表示该地中度适生; “EI≥30”表示该地高度适生。
1.3 利用GARP进行适生性分析 1.3.1 资料来源从http://www.nhm.ku.edu/desktopgarp/网站下载Desktop GARP(Version 1.1.6)软件以及各大洲的环境数据层,每一环境层均包含年平均温度、年最高温度、年最低温度、年降水量、地貌、数字高程、坡度、植被、灌溉水累积量、灌溉水流向、年蒸发量、年湿度、年辐射、年有霜期等14个环境因子图层,图层栅格分辨率为0.1°。从国家基础地理信息系统(http://nfgis.nsdi.gov.cn/)下载获取1 : 400万的中国行政区划图作为分析底图。
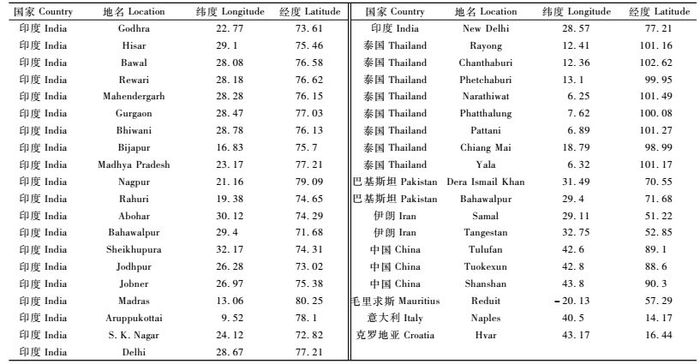
1.3.2 方法与步骤1) 枣实蝇地理分布数据收集与整理整理获得枣实蝇印度分布点21个(Arora et al., 1999; Basha,1952; Batra,1953; Bagle,1992; Dashad et al., 1999; Khare,1923; Pareek et al., 2003; Singh et al., 2002),泰国分布点8个(Hancock et al., 1994),巴基斯坦分布点2个(Bashir et al., 2005; Stonehouse et al., 1998),伊朗分布点2个(Farrar et al., 2004),毛里求斯分布点1个(Anon,1987),意大利分布点1个(Costa,1854),克罗地亚分布点1个(Frauenfeld,1867),结合我国3个分布点(阿地力·沙塔尔等,2008),一共收集整理得到39个枣实蝇已知分布点,按GARP要求制成经纬度坐标数据EXCEL文件。
2) 参数设置的确定设置参数时,选择20个分布点作为训练数据,余下的作为验证数据,调试其他参数,预测枣实蝇在全世界的分布地点。利用已知分布点对预测结果进行比较,直到预测结果准确为止。据此从算法(如生物气候规则或逻辑斯谛回归)中归纳出一种参数设置方法(Stockwell et al., 1992; Stockwell,1999),再运用已知分布点,预测亚洲地区枣实蝇的分布区域。
3) 气候适生程度划分预测结果以ARC/INFOGRID数据格式输入ArcGIS,利用空间分析模块来考察模型间的一致性。根据Anderson等(2003)提供的方法将运算得到的10个最优模型进行叠加,计算模型中的每个栅格的适生值(对于单个模型,1代表预测适生,0代表预测非适生)。在叠加后的图层中,每个栅格的适生值等于预测该栅格为适生区的模型数量(Anderson et al., 2002)。本研究将适生值大于3的划分为适生区,其中3 ~ 5为一般适生区,5 ~ 7为中度适生区,7 ~ 10为高度适生区。预测结果利用ArcGIS软件进行反距离加权插值处理,得到枣实蝇在我国的气候适生区预测图。
1.4 枣实蝇在我国各省市区总体适生程度判定参考陈洪俊等(2007)的方法,本文以CLIMEX和GARP 2种软件的适生性预测结果的重叠部分作为枣实蝇在我国的主要气候适生区,非重叠部分为次要气候适生区域; 其他区域则为非适生区。
枣实蝇潜在适生区必须满足气候适生和有枣树分布2个条件,否则为非潜在适生区。由于枣树在我国的分布资料只收集到省级水平,没有具体到各县市,因此只在省市区尺度上评价枣实蝇适生程度,并划分为高、中、低3个等级。划分时的假设前提,即当物种能适应该地区生态气候条件时,无论适生程度的高低,决定物种能否生存发展的关键在于寄主的丰富程度。因此最终各省市区枣实蝇适生程度的高低主要由寄主的种植密度决定。
1.5 枣实蝇对我国各省市区危险性的定量评估根据枣实蝇生物生态学特性、可能造成的危害及其在我国各适生区的发生情况,参照国际上有害生物危险性分析方法,采用“多指标综合评估方法”(蒋青等,1995; 黄振等,2008),从分布(P1)、潜在危害性(P2)、受害寄主的经济重要性(P3)、传入可能性(P4)和风险管理的难易程度(P5)等5个方面建立了枣实蝇PRA评估量化分析指标评判标准。各指标依据评判标准,在听取专家意见后,按3,2,1,0等4个等级打分,对枣实蝇在各省市区的危险性进行评估。5个方面的具体评判指标、标准及分值(用括弧表示)如下。
评判地区内分布情况P1 :区内无分布(3),分布面积占0 ~ 20% (2),占20% ~ 50% (1),大于50% (0)。P2具有3个指标,潜在危害性P21 :被害株死亡率(产量损失)在20%以上(3),在20% ~5% (2),在5% ~ 1% (1),在1%以下(0) (影响品质者对应地再加1,但最高值为3);是否为其他检疫性有害生物的传播媒介P22 :可传带3种以上的检疫性有害生物(3),传带2种(2),传带1种(1),不传带(0);国外重视程度P23 :有20个以上的国家把枣实蝇列为(补充)检疫性有害生物(3),19 ~ 10个(2),9 ~ 1个(l),无(0)。P3具有3个指标,受害寄主种类P31 : 10种以上(3),9 ~ 5种(2),4 ~ 1种(1),无寄主(0);受害寄主面积比例P32 :大于1 /300(3),1 /300 ~ 1 /900 (2),1 /900以下(1),0 (0);受害寄主特殊经济价值P33 :根据其应用价值、出口创汇等方面,进行判断定级(P33 = 3,2,1,0)。P4具有5个指标,评判地区外分布情况P41 :评判地区外50%以上国家/省有分布(3),在50% ~ 25%之间(2),25% ~ 0 (1),无分布(0) (临近评判地区有分布再加1,但最高值为3);截获的难易程度P42 :经常被截获(3),偶尔被截获(2),从未被截获(1),因现有检疫技术的原因本项不设0级; 运输中存活率P43 : 40%以上(3),40% ~ 10% (2),10%以下(1),0(0);传播能力P44 :气传(3),由活动能力很强的媒介传播(2),土传或活动能力很弱的媒介传播(1),该项不设0级; 评判地区内适生范围P45 : 50%以上区域适生(3),50% ~ 25% (2),25%以下(1),无适生区(0)。P5具有3个指标,检疫鉴定难度P51 :现有方法可靠性低则费时(3),较可靠但费时(2),可靠但技术要求高、难度大(1),非常可靠、简便、快捷(0);除害难度P52 :现有除害处理技术几乎完全不能杀死枣实蝇(3),除害率50%以下(2),除害率50% ~ 100% (1),除害率100% (0);根除难度P53 :田间防治效果差、成本高、难度大(3),效果显著、成本很低、简便(0),介于上述两者之间(P53 = 2,1)。
对各评判指标进行赋值后,按照有害生物危险性定量分析计算公式(蒋青等,1995; 黄振等,2008),计算出危险性总R值。具体公式为:P2 =0.6P21 + 0.2P22 + 0.2P23; P3 = Max(P31,P32,P33); 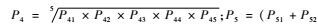
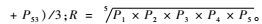
我国林业有害生物的危险程度一般划分为4级,在参考相关文献(吴海军等,2007)及听取专家意见的基础之上,本文将枣实蝇在我国的风险值等级做出如下划分,2.0<R≤3.0为高度风险,1.0<R≤2.0为中度风险,0<R≤1.0为低度风险,R = 0为无风险。
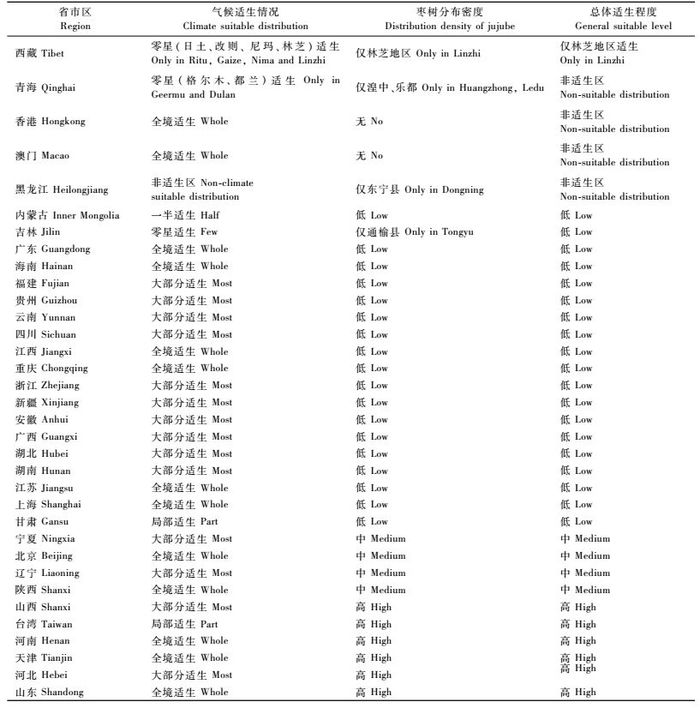
2 结果与分析 2.1 枣树在我国的分布概况中国种植信息网中无西藏、青海、内蒙古、吉林、黑龙江、广东、海南、浙江、台湾、香港、澳门等地的枣产量资料。查阅其他资料获知,以上未统计到的省区中除香港、澳门未能收集到枣树分布资料外(预测时假设2地无枣树分布),均有枣树分布。其中浙江为中国南方枣树栽培中心,1993—1995年全省枣产量平均为3 900 t(刘孟军,2000); 台湾省共有青枣2 000 hm2,产量高达9万kg·hm-2 (http://www.huaxia.com/la/tbchwz/2005/00320151.html)。其余省区虽有枣树分布,但种植均不成规模,因此预测时,假设这些省区均为枣树低密度种植区。具体如下:西藏仅林芝地区产红枣(李健等,2003); 青海省在东北部湟水谷地、海拔2 000 m左右的乐都(http://www.qhda.gov.cn/readnews.asp?newsid=437)、循化(马秀清等,2007)有红枣栽培以及酸枣的分布; 内蒙古枣树主要分布在东西部小气候较好的山麓地带,如赤峰的宁城、喀拉沁旗、呼和浩特至包头的沿山南麓一带,伊克昭盟的马珊,准格尔旗等地(http://www.hongzaowang.com/shownews.asp?newsid=3119); “寒脆枣”在吉林省西部通榆选育成功,填补了其枣树栽培的空白(http://sr.yuanlin.com/html/srnews/detail/2006-12/5333.html); 2005年黑龙江东宁县粮种场栽培枣树成功,结束了黑龙江省无枣树栽培的历史(http://news.xinhuanet.com/newscenter/2005-06/22/content_3120846.htm); 南亚热带作物研究所的科技人员将毛叶枣(Z.mauritiana)成功引入广东、海南和广西(http://stcncd-liuxian.gdcct.gov.cn/news/2005/2005-12-05/1133773124.html)。枣树在我国各省市区的种植情况,如表 1所示。
|
|

表 1中需要注意的是:由于评定结果为相对省区面积产量,因此某些枣产量较高的省份,其最终种植密度评定结果较低,比如新疆和甘肃两省区。另由表 1知:我国枣果主产区为东中部地区的陕西、山西、河南、天津、北京、河北和山东等7个省市,2007年上述7个省市枣总产量为256.413 0万吨,占全国总产量的85%。
2.2 枣实蝇在中国的适生性分析 2.2.1 CLIMEX中枣实蝇生物学参数设定根据国外,特别是印度、泰国等已知枣实蝇分布点进行参数调试,当CLIMEX预测结果与实际分布达到最大的吻合后,确定此时的参数为枣实蝇最佳参数,如表 2所示。
|
|
另外根据Lakra等(1985)的研究,结合在吐鲁番地区的实际调查情况,在CLIMEX3.0软件中设置了灌溉措施,选择按月实施(并选择“灌溉总量包括自然降雨和人为浇水”一项),从4—9月灌溉系数分别为0.3,1.4,2.8,2.8,1.4,0.2 mm·d-1。
2.2.2 ClIMEX预测结果分析CLIMEX计算结果发现:在枣实蝇源发地印度的43个气象站点中,EI值大于0的气象站点一共有41个,占总数的95%,EI值大于20的站点约占全国的70%;在泰国和伊朗,全国EI平均值分别为32和35,均为高度适生; 意大利那不勒斯EI值为39;克罗地亚Hvra岛EI值为37;我国枣实蝇发生地吐鲁番市、鄯善县和托克逊县的EI值分别为15,29和27,适生程度中等,这与实际情况十分吻合。
CLIMEX计算结果显示:黑龙江、青海、西藏、台湾、上海等5个省市所有站点的EI值均为0。EI值不为0即适生站点的总数为390个,占站点总数的53%。其中中度适生程度“10≤ EI<30”的站点数为202个,占适生站点总数的52%;高度适生程度“EI≥30”的站点数为91个,占适生站点总数的23%;中高度适生程度站点共计占适生站点总数的75%。由此可知,枣实蝇在我国的适生范围较广、适生程度中等。
利用ArcGIS软件空间分析中的反距离加权法对EI值进行插值分析,得到枣实蝇在我国的生态气候适生区分布(图版Ⅰ- 1) :枣实蝇在我国潜在适生范围南至海南岛、北至新疆北纬47度地区。核心适生区位于陕西、山西、河北、北京、天津、河南和山东7省市的大部分地区。其他气候适生区包括:新疆绝大部分地区; 甘肃同新疆邻近的安西、敦煌一带,同内蒙古西南相接的民勤、鼎新、金塔一带,同宁夏西南相邻的靖远一带,同陕西、四川相接的天水、武都; 内蒙古西部地区; 辽宁; 吉林的集安和白城; 内蒙古东部与辽宁、吉林接壤局部地区; 江苏绝大部分地区; 安徽绝大部分地区; 浙江西部局部地区; 福建; 江西; 西藏同新疆接壤的北部局部地区; 四川中东部广大地区; 重庆; 贵州; 云南; 湖北绝大部分地区; 湖南; 广西; 广东; 香港; 海南; 台湾西部局部地区。

|
图版Ⅰ Plate1 1.CLIMEX3.0软件预测枣实蝇在中国的生态气候适生分布区;2.GARP预测枣实蝇在我国的适生区分布区;3.CLIMEX和GARP预测结果叠加1.The potential distribution of C.vesuviana in China predicted by CLIMEX3.0; 2.The potential distribution of C.vesuviana in China predicted by GARP; 3.The superposition chart of the results of CLIMEX and GARP. |
枣实蝇在全球分布点的经纬度坐标见表 3。
|
|
经ArcGIS软件的处理,得出枣实蝇在我国的气候适生区预测图,具体如图版Ⅰ- 2所示。结果显示枣实蝇在我国的气候适生区分布广泛,分布北界达到了新疆阿勒泰市和布尔津县南部,向南则分布于整个海南省。
由图版Ⅰ- 2知:GARP预测枣实蝇在我国的核心气候适生区域位于河北、北京、天津、河南、山东、江苏、上海市和安徽等8个省市,其余适生区分布于:新疆绝大多数地区; 甘肃与新疆、内蒙古和宁夏接壤的零星地区; 青海格尔木市、都兰县2地; 宁夏北部; 陕西北部地区及南部零星地区; 内蒙古西部地区、东部与吉林、辽宁接壤的地区; 山西南部; 辽宁大部分地区; 吉林靠近内蒙古和辽宁的局部地区; 浙江北部的零星地区; 江西和福建的绝大部分地区; 湖北和湖南2省的东部地区; 广西和广东2省区的大部分地区; 海南全省; 西藏墨脱、察隅、错那3县南部; 云南中南部; 四川东南部; 重庆与四川相接的西部局部地区; 贵州与云南和广西相接的局部地区; 台湾西南部。
2.4 CLIMEX和GARP预测结果比较与分析将CLIMEX和GARP分别得到的预测结果进行叠加,其结果如图版Ⅰ- 3所示。比较发现,两者预测枣实蝇在我国的气候适生区域范围总体一致,均认为枣实蝇在我国具有广泛的适生范围,从内陆地区到沿海地区均有枣实蝇潜在气候适生区,分布的北界到达新疆北纬47°的地区,向南达到海南岛的最南端。具体存在以下异同点:
1) 两者一致认为黑龙江不是枣实蝇适生区,对吉林、青海、西藏3省区的预测结果总体趋于一致,只是CLIMEX结果经插值后认为吉林省集安市,青海省北部的海西蒙古自治州,西藏北部的日土、改则、尼玛3县的北部地区,西藏东部与四川交界的贡觉、芒康、盐井3县为枣实蝇适生区; 而GARP认为吉林靠近内蒙古和辽宁的局部地区,青海省格尔木市和都兰县2地,西藏墨脱、察隅、错那3县南部为枣实蝇适生区。
2) 两者预测枣实蝇在我国的气候适生范围基本一致,但具体地区有差异。对于新疆、四川、贵州、重庆的预测结果,CLIMEX预测结果较GARP广泛。CLIMEX认为枣实蝇中心适生区域位于陕西、山西、河北、北京、天津、河南和山东等7省市,GARP则认为枣实蝇在我国的中心适生区域位于河北、北京、天津、河南、山东、江苏、上海和安徽等8省市。后者的预测结果更偏向我国的东部地区。具体如图 3所示(深色部分为重叠区域)。
综合两者预测结果,枣实蝇在我国的主要气候适生区域为二者预测结果的重叠部分,主要位于全国的23个省市区,即:新疆环塔里木盆地的南疆各地州、哈密地区、吐鲁番地区和昌吉州; 甘肃与新疆、内蒙古和宁夏接壤的零星地区; 内蒙古西部; 内蒙古东部与吉林、辽宁接壤的部分地区; 宁夏北部地区; 四川东南部地区; 云南中南部地区; 陕西中北部地区; 山西南部地区; 河北除北部局部地区外的绝大部分地区; 北京; 天津; 河南除西部局部地区外的绝大部分地区; 辽宁中西部地区; 山东; 江苏与安徽除南部局部地区外的绝大部分地区; 湖北中东部地区; 湖南中部局部地区; 江西绝大部分地区; 福建大部分地区; 广西与广东除北部局部地区外的绝大部分地区; 海南省。次要适生区域主要位于新疆北部和南部的局部地区; 内蒙古临河以东局部地区; 宁夏南部局部区域; 甘肃与新疆相接的局部地区; 甘肃与四川、陕西三省交界的局部地区; 青海格尔木市、都兰县; 陕西南部地区; 山西北部地区; 河北北部地区; 吉林与内蒙古、辽宁接壤的局部地区; 四川中部; 重庆东部; 西藏区墨脱、察隅、错那3县南部; 湖北与湖南的西部地区; 贵州北部地区; 浙江北部局部地区; 广西与广东的北部局部地区; 台湾西部地区。
结合枣树在我国的种植概况,对枣实蝇在我国各省市区内的总体适生程度进行评价,分析认为河北(包括北京和天津)、河南、山东、山西、陕西、台湾、宁夏、辽宁等主要枣产区为枣实蝇在我国的中高度适生区域,青海、香港、澳门、黑龙江等省市区为枣实蝇非适生区,西藏仅林芝地区为枣实蝇适生区,其余省市区由于枣树种植密度较低,均为枣实蝇低度适生区。具体结果见表 4。
|
|
按照有害生物定量评价指标体系的赋值规则,在听取专家意见后,对各省市区内的相关指标进行打分,根据公式计算出危险系数R值,其具体结果如表 5所示。
|
|
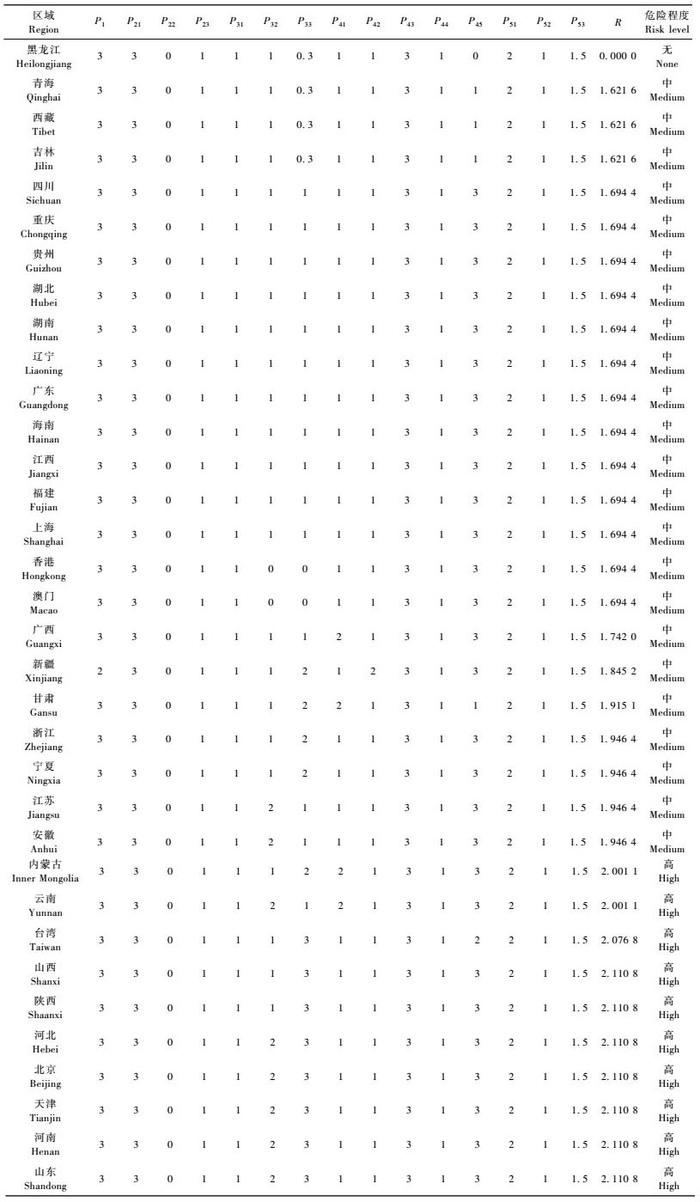
由表 5知:全国除黑龙江省不受枣实蝇威胁外,其余各省市区均受到枣实蝇的潜在威胁,危险程度为中高度,且在我国枣果的5大主产区内危险程度均为高度,由此预见枣实蝇对我国枣产业的发展构成巨大的潜在威胁。
3 讨论运用CLIMEX软件进行预测时,所选气象站点的数量及覆盖范围将影响预测结果的准确度,往往由于气象站点数据不够而影响整体预测结果。如台湾省只有台北和花莲2个气象点,其预测结果为此2个站点EI值为0,即为非适生站点,进而整个台湾省被预测为非适生区,但GARP预测认为整个台湾西部均为枣实蝇气候适生区; 西藏北部也由于无气象站点记录,而被预测为非适生区,但利用ArcGIS插值处理发现西藏北部为枣实蝇潜在适生区。因此在利用CLIMEX进行预测时,气象站点数据获取是重要基础。另外利用ArcGIS中的插值功能,可以解决各站点之间相互独立的问题。
本文利用了CLIMEX和GARP两种软件对枣实蝇适生区进行预测,一则因为枣实蝇相关资料较少,另外还因为2种软件在预测上具有互补特性。GARP运用遗传算法,分析物种已知生存地区的温度、湿度、海拔、降水等14个环境因子,推算出适合物种生存的最佳环境条件; CLIMEX根据物种生长发育的生物学数据,分析影响种群建立的环境因子来分析环境的适合度,对潜在区域做出判断,得出预测结果。2种软件预测时切入点不同,前者注重环境因子,后者注重物种自身生物学特性,因此两者预测结果并不冲突而互为补充。
枣实蝇危险性的综合评估表明:该虫抗逆性强,生存繁殖率高,对枣树的危害程度严重,造成的损失极大,传入后扩散快,根除困难,是高度危险的检疫性有害生物,特提出以下建议:
进一步加强检疫监管力度,制定相关的检疫法规,禁止疫区内相关枣产品向疫区外销售,制止单位或个人从疫区偷调枣产品的行为,防止枣实蝇进一步扩散蔓延。
对从枣实蝇发生国家特别是与新疆接壤的吉尔吉斯坦、塔吉克斯坦、乌兹别克斯坦、阿富汗、巴基斯坦,与云南、广西邻近的泰国等国家进口的枣产品如鲜枣、干枣,实施严格的检疫,防止枣实蝇从境外传入我国其他省份; 运输新鲜枣果经过吐鲁番疫区时,停留一律不得超过24 h。
在新疆其他地区以及全国其他枣树种植省份宣传普及枣实蝇相关知识,大力开展枣实蝇调查与监测工作,加强民间尤其是枣农的防范意识;
吐鲁番地区非疫区内加强枣实蝇监测和预防工作,尤其是托克逊县非疫区与疫区之间的缓冲区内枣实蝇的监测和预防工作。
疫区内综合采用化学、物理等措施对枣实蝇采取全部种群治理,彻底消灭枣实蝇。
疫区内选育抗虫性强、品质优良的枣树品种,提高农林复合生态系统自身抵御能力。
阿地力·沙塔尔, 何善勇, 田呈明, 等. 2008. 枣实蝇在吐鲁番地区的发生及蛹的分布规律[J]. 植物检疫, 22(5): 295-297. |
陈洪俊, 刘海军, 李镇宇, 等. 2007. 西花蓟马在我国的潜在分布区预测[J]. 植物检疫, 3(21): 160-164. |
何善勇, 朱银飞, 阿地力.沙塔尔, 等. 2009a. 枣实蝇蛹发育起点温度和有效积温测定[J]. 昆虫知识, 46(5): 935-938. |
何善勇, 朱银飞, 阿地力.沙塔尔, 等. 2009b. 吐鲁番地区枣实蝇发生规律[J]. 昆虫知识, 46(6): 930-934. |
黄振, 黄可辉. 2008. 桔小实蝇传入中国风险的定量分析[J]. 江西农业学报, 20(8): 61-62. |
蒋青, 梁忆冰, 王乃杨, 等. 1995. 有害生物危险性评价的定量分析方法研究[J]. 植物检疫, 9(4): 208-211. |
刘孟军. 2000. 国内外枣树生产现状、存在问题和建议[J]. 中国农业科技导报, 2(2): 76-79. |
李健, 郑宇, 郑惠章. 2003. 西藏林芝地区果树资源开发利用规划设想[J]. 柑桔与亚热带果树信息, 19(10): 1-3. DOI:10.3969/j.issn.1673-1514.2003.10.001 |
马秀清, 马光辉, 韩学镇. 2007. 循化枣树的栽培技术[J]. 青海农林科技, (4): 85-86. |
周国梁, 陈晨, 叶军, 等. 2007. 利用GARP生态位模型预测桔小实蝇(Bactrocera dorsalis)在中国的适生区域[J]. 生态学报, 27(8): 3362-3368. |
吕文刚, 林伟, 李志红, 等. 2008. 枣实蝇在中国适生性初步研究[J]. 植物检疫, 22(6): 343-347. |
吴海军, 李友莲, 丁三寅, 等. 2007. 入侵生物苹果绵蚜在中国的风险性分析[J]. 山西农业大学学报:自然科学版, 27(4): 368-371. |
Anderson R P, Gómez-Laverde M, Peterson A T. 2002. Geographical distributions of spiny pocket mice in South America: insights from predictive models[J]. Global Ecology and Biogeography, 11(2): 131-141. DOI:10.1046/j.1466-822X.2002.00275.x |
Anderson R P, Lew D, Peterson A T. 2003. Evaluating predictive models of species ' distributions: criteria for selecting optimal models[J]. Ecological Modelling, 162(3): 211-232. DOI:10.1016/S0304-3800(02)00349-6 |
Anon. 1987. Annual Report, Entomology Division, Ministry of Agriculture, Fisheries and Natural Resources, Reduit, Mauritius.
|
Arora P K, Nirmal K, Batra R C, et al. 1999. Physico-chemical characteristics of some ber varieties in relation to fruit fly incidence[J]. India Journal of Appllied Horticulture, 1(2): 101-102. |
Bagle B G. 1992. Incidence and control of fruit fly Carpomyia vesuviana Costa of ber Zizyphus mauritiana, Lamk[J]. Indian Journal of Plant Protection, 20: 205-207. |
Basha J M G. 1952. Experiments on the control of the fruit borers of jujube(Zizyphus spp.) Carpomyia vesuviana Costa and Meridarchis scyrodes Meyr in the South India[J]. Indian J Entomol, 14: 229-238. |
Batra H N. 1953. Biology and control of Dacus diversus Coquillent and Carpomyia vesuviana Costa and important notes on other fruit flies in India[J]. Indian J Agric Sci, 23: 87-112. |
Bashir A, Rukhsana A, Amir A, et al. 2005. Comparison of different methods to control fruit fly (Carpomyia vesuviana) on Ber (Zizyphus mauritiana)[J]. Pakistan Entomology, 2(27): 1-2. |
Costa A. 1854. Frammenti di entomologia napoletana. Annali Scientifici (Napoli) 1: 69-91.
|
Dashad S S, Chaudhary O P, Rakesh O. 1999. Studies on the incidence of ber fruitfly (Carpomyia vesuviana Costa) in south-western Haryana[J]. Crop Research (Hisar), 18(1): 115-118. |
Farrar N, Asadi G H, Golestaneh S R. 2004. Damage and host ranges of Ber Fruit fly Carpomyia vesuviana Costa (Tephritidae) and its rate of parasitism[J]. Agricultural Sciences, Faculty of Agriculture, University of Guilan, 5(1): 120-130. |
Frauenfeld G R von. 1867. Das Insektenleben zur See und zur Fauna und Flora von Neucaledonien etc[J]. Verh Zool Bot Ges Wien, 17: 425-502. |
Hancock D L, Drew R A I. 1994. New species and records of Asian Trypetinae (Dipetera: Tephritidae)[J]. Raffles Bulletin of Zoology, 42(3): 1-37. |
Khare J L. 1923. Ber (Zizyphus jujube) fruit and its fly pest[J]. Agic Res Inst Pusa Bull, 143: 1-16. |
Lakra R K, Singh Z. 1983. Oviposition behavior of ber fruitfly Carpomyia vesuviana and relationship between its incidence and ruggedness in fruits in Haryana India[J]. Indian Journal of Entomology, 45: 48-59. |
Lakra R K, Singh Z. 1985. Seasonal fluctuations in incidence of ber fruitfly Carpomyia vesuviana Costa (Diptera: Tephritidae) under agro-climatic conditions of Hisar[J]. Haryana Agric Univ J Res, 15(1): 42-50. |
Pareek S, Fagera M S, Dhaka R S. 2003. Genetic variability and association analysis for fruitfly (Carpomyia vesuviana Costa) infestation in ber[J]. Indian Journal of Plant Protection, 31(2): 89-90. |
Sangwan M S, Lakra R K. 1992. Effect of temperature and soil depth levels on pupae of jujube fruit fly, Carpomyia vesuviana Costa[J]. Journal of Insect Science, 5(1): 80-81. |
Singh M P, Vashishtha B B. 2002. Factors influencing infestation of fruit-fly (Carpomyia vesuviana) in indian jujube[J]. Indian Journal of Agricultural Sciences, 72(9): 543-547. |
Stockwell D R B, Noble I R. 1992. Induction of Sets of Rules From Animal Distribution Data:A Robust and Informative Method of DataAnalysis[J]. Mathematics and Computers in Simulation, 33(526): 385-390. |
Stockwell D R B. 1999. The GARP modelling system: problems and solutions to automated spatial prediction[J]. International Journal of Geographical Information System, 13(2): 143-158. DOI:10.1080/136588199241391 |
Stonehouse J M, Mumford J D, Mustafa G. 1998. Economic losses to Tephritid fruit flies (Diptera: Tephritidae) in Pakistan[J]. Crop Protection, 17: 159-164. DOI:10.1016/S0261-2194(97)00091-4 |
Sutherst R W, Maywald G F, Yonow T, et al. 1999. CLIMEX: Predicting the effects of climate on plants and animals. Melbourne: CSIRO.
|