 2011, Vol. 47
2011, Vol. 47文章信息
- 刘文飞, 樊后保
- Liu Wenfei, Fan Houbao
- 杉木人工林凋落物C,N,P归还量对氮沉降的响应
- Impacts of Nitrogen Deposition on C, N, and P Fluxes in the Litterfall of Chinese fir Plantation
- 林业科学, 2011, 47(3): 89-95.
- Scientia Silvae Sinicae, 2011, 47(3): 89-95.
-
文章历史
- 收稿日期:2009-09-07
- 修回日期:2010-12-07
-
作者相关文章



预测表明,人类活动使得进入陆地生态系统的无机氮至少增加1倍,而且由于人口增长、化石燃料消费的增加以及日益增加的对工业化肥的依赖,未来几十年内氮的输入将会持续增加(Galloway et al., 1995; Vitousek et al., 1997)。1860年全球人类每年所产生的活性氮(Nr)约15 Tg N,到20世纪90年代初期增加到156 Tg N,2050年预计将达到270 Tg N; 大气NOx和NH3的排放总量也从1860年的23Tg N增加到20世纪90年代初期的93 Tg N,2050年将达到189 Tg N(Galloway et al., 2004)。因此,陆地生态系统氮超负荷的问题已引起科学家和公众的广泛关注(Nihlgard,1985; Kochy et al., 2001)。
全球森林生态系统的碳储存和碳循环正在受到氮沉降的深刻影响,因为氮沉降改变了陆地生态系统的生产力和生物量积累(樊后保等,2007)。然而,关于氮沉降能否增加森林生态系统的碳储存,在科学界还存在激烈的争论。按照合理的推测,在氮素受限的生态系统中,从大气沉降中增加的有效氮供应可能会导致生物量生产的增加,其结果是增加额外的碳固定量(Vejre et al., 2003)。然而,其他的预测结果表明,氮素对生态系统的刺激作用不可能引起明显的碳贮存(Korner,2000),而实际上是减少生态系统生产力和碳储量(Schulze,1989; Cao et al., 1998)。森林是陆地生态系统中最大的有机碳库,而森林凋落物作为森林生态系统碳库的重要组成部分在陆地生态系统碳循环中起着重要的作用(Christopher,1999)。据估计,全球每年通过凋落物分解归还到土壤的有机碳约为50 Gt(Palviainen et al., 2004),凋落物的生产与分解及其相关过程变化直接影响着陆地碳储量和大气二氧化碳浓度(Fang et al., 2006)。
到目前为止,国内外对氮沉降与凋落物分解的研究较多(樊后保等,2008; Micks,2004; Prescott,2004),但有关凋落物量及其养分归还量影响的研究鲜见报道(樊后保等,2007)。之所以选择杉木(Cunninghamia lanceolata)作为研究对象,主要是因为杉木是我国南方重要的用材树种之一,自然分布和人工栽培都很广,是中国亚热带地区典型的森林生态系统。其次,以往的研究表明,对于氮沉降针叶林林较阔叶敏感。因此,在氮沉降全球化的背景下,开展杉木人工林凋落物C,N,P方面的研究,为全球变化下的碳库和碳循环研究提供理论支持,为我国进一步开展氮沉降研究打好基础,同时为杉木林的可持续经营提供理论指导。
1 材料和方法 1.1 试验地概况与样地设置试验地设在福建三明沙县官庄国有林场,位于117°43′29″ E,北纬26°30′47″ N。属中亚热带季风气候,四季气温冷暖适中,日照充足,年平均气温18.8 ~19.6 ℃,年平均降水量1 606 ~ 1 650 mm,无霜期271天。试验林设置在该林场的白溪工区21林班8小班南坡上,平均海拔200 m左右,土壤为山地红壤。试验林为1992年营造的杉木人工林,面积为6 hm2。林下植被稀疏,以五节芒(Miscanthus floridulus)、芒萁(Dicranopteris olichotoma)、蕨(Pteridium aquilinum var.latiusculum)等为主,盖度在3% ~ 5%。选择立地和林分条件相似的杉木人工林建立12块固定试验样地,样地设置按完全随机区组设计,样地面积均为20 m × 20 m,内设15 m × 15 m中心区域,以便破坏性试验在中心区外围的处理区进行。2003年12月进行本底值调查,结果见表 1。
|
|
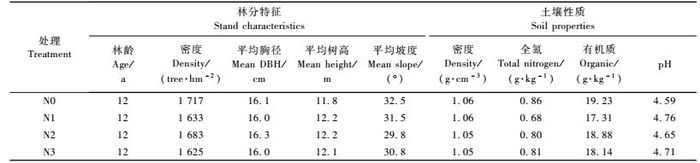
我国亚热带地区氮沉降分布存在很强的地域差异,福建省南部平均降水氮输入量为53 kg N·hm-2 a-1 (肖健,2005),中西部地区降水氮输入量只有11.4 ~ 18.1 kg N·hm-2 a-1 (樊后保等,2000); 广东省鼎湖山自然保护区1989—1990年和1998—1999年的降水氮输入量分别为35.57和38.14 kg N·hm-2 a-1 (黄忠良等,1994; 周国逸等,2001); 而在江西省氮沉降向林地输入的氮达到82.8kg·N hm-2 a-1 (樊建凌等,2007)。同时,我国氮沉降从总体上有不断上升的趋势。考虑到这种时空变化特征,设计较大梯度模拟氮沉降水平能更好地反映整个亚热带地区的氮沉降分布状况和增长趋势。因此,按氮施用量的高低,分4种处理,从高到低分别以N0(对照,0 kg N·hm-2 a-1),N1(60 kg N·hm-2 a-1),N2(120 kg N·hm-2 a-1),N3 (240 kg N·hm-2 a-1)表示,每个处理均设3个重复。2003年12月建立样地后,于2004年1月开始进行模拟氮沉降处理,在整个试验期间,每月月初以溶液的形式给样地喷洒。按照处理水平的要求,将每个样方每次所需要喷施的CO(NH2)2溶解在20 L水中后,以背负式喷雾器在林地人工来回均匀喷洒。对照样方喷施同样量的水,以减少因外加的水而造成对森林生物地球化学循环的影响。
1.2.2 凋落物收集方法在各样地内随机设置10个1 m × 1 m的凋落物收集器,定期(每月底)收集落在收集器上的凋落物,装入塑料袋带回实验室,区分别落叶、落枝、树皮、落果(花、果实、种子等)及其他碎屑物(包括昆虫残体与粪便及鸟类粪便等)等组分,在80 ℃恒温条件下烘干48 h后称量。收集时间从2004年1月开始,本文选择模拟氮沉降2年后2006年全年的凋落物样品。
1.2.3 凋落物C,N和P的测定方法凋落物各组分粉碎后,全年样品按1—3月、4—6月、7—9月和10—12月4个季度进行分析,测定时,如1—3月凋落物叶,称取1,2,3月凋落物叶样品各1 g,3个月充分混合,以此类推,将混合均匀的样品精确称量1 ~ 2 mg,由元素分析仪(美国PE公司-2400II型)测得各组分C,N含量,钼锑抗法(美国PE公司-Lambda25型紫外分光光度计)测定各组分P含量。
1.2.4 数据分析利用DPS软件进行统计分析,然后以LSD多重检验法检验不同处理间的差异显著性。
2 结果与分析 2.1 凋落物量对氮沉降的响应由表 2可以看出,经过不同的氮沉降处理(N0,N1,N2,N3)后,杉木人工林的年凋落量分别为1 008.83,1 164.10,1 147.30和976.47 kg·hm-2。分析显示,低氮(N1)和中氮(N2)在一定程度上增加森林凋落物量,而高氮处理(N3)则表现出一定的抑制作用,N3与N1,N2处理凋落物量差异达到显著水平(P<0.05)。
|
|
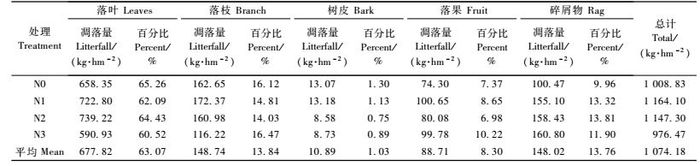
凋落物的组成中(表 2),落叶占总凋落量的60.52% ~ 65.26%,其他依次为落枝(13.84% ~16.47%)、碎屑物(4.98% ~ 7.70%)、落果(6.98% ~ 10.22%)和树皮(0.75% ~ 1.30%)。
2.2 不同N处理凋落物各组分C,N和P年平均含量的比较从表 3可知,各组分中C,N和P元素含量大小顺序为C>N>P,但不同N处理各元素含量不尽相同,以落叶为例:落叶中,C和N元素含量随氮沉降水平的增加而增加,氮沉降表现出增大凋落物叶C,N元素含量的作用,经N1,N2和N3处理凋落叶中C,N元素含量比N0处理分别提高0.94%,4.11%;0.97%,6.85%; 2.51%,27.40%。N3与N0,N1和N2处理凋落叶中N元素含量达到显著水平,但C元素含量差异不显著。P元素含量大小顺序为N2 >N1>N0>N3,说明N1,N2处理能提高凋落叶P含量,而N3表现为抑制作用,但各处理差异没有达到显著水平。
|
|

在已知凋落量及其养分含量的前提下,就可以计算凋落物的养分归还量,结果见表 4。不同N处理凋落物各组分中元素归还量的大小顺序为C >N>P。N0,N1,N2和N3处理凋落物C元素的归还量分别为474.70,544.07,538.55和474.02kg·hm-2,相对于N0处理,N1,N2处理显著提高凋落物C元素的归还量(P<0.05),N3则表现为抑制作用,但差异没有达到显著水平。
|
|
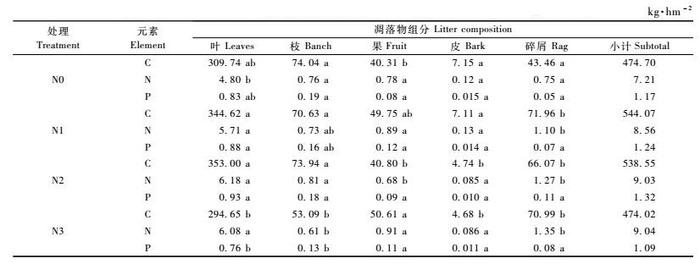
N元素的归还量分别为7.21,8.56,9.03和9.04 kg·hm-2,经N1,N2和N3处理后,凋落物N元素归还量分别比N0处理高出18.72%,25.24%和25.38%,并且与N0处理差异均达到显著水平(P<0.05)。
P元素的归还量分别为1.17,1.24,1.32和1.09 kg·hm-2,N1,N2处理表现为提高凋落物P元素归还量的作用,N3则表现为抑制作用,N3处理与N1,N2处理差异达到显著水平(P<0.05)。
2.4 凋落物中C,N和P元素归还量的时间动态由图 1可知,各N处理凋落物C,N和P元素在年中均出现2个比较明显的归还高峰期,第1次峰值均出现在4月,第2次峰值除N0处理凋落物C,P元素的归还量和N1,N2处理凋落物的P元素的归还量出现在7月,其余均出现在6月。

|
图 1 各处理凋落物元素归还月动态变化 Figure 1 Monthly dynamic of elements flux in litter under different nitrogen treatments |
各N处理凋落物C,N和P元素归还的极值均出现在4月,该月份C元素归还量分别占全年归还量的22.78%,29.48%,23.43%和21.83%,N1处理与N0,N3处理差异达到显著水平(P<0.05);该月份N元素归还量占全年归还的22.94%,29.14%,25.54%和22.02%,N0处理与N1,N3处理差异达到显著水平(P<0.05);该月份P元素归还量占全年归还量的20.98%,24.87%,21.03%和18.68%,N1处理与N3处理差异达到显著水平(P<0.05)。
2.5 氮沉降下凋落物各组分C/N比值变化由图 2可以看出,各氮处理C/N比值的趋势和不同凋落物组分对氮沉降的响应有所不同,除N1处理的落叶和落果的C/N比值在1年中表现为先下降后上升外,其余处理C/N比值均表现为下降-上升-下降。

|
图 2 各处理凋落物各组分C/N比值月动态变化 Figure 2 Monthly dynamic of C/N ratio in litter composition under different nitrogen treatments |
以落叶为例,N0处理的C/N比值在1年中的变化范围为59.25 ~ 79.29,N1,N2和N3处理分别为57.15 ~ 69.71,50.20 ~ 68.90和43.70 ~ 61.91,说明经N1,N2和N3处理后,落叶C/N比值的平均值比N0处理分别下降0.27%,5.78%和17.98%,并且N3处理与N0,N2处理差异达到显著水平(P<0.05),这也反映在经过不同水平的模拟氮沉降处理后,凋落叶中N含量的提高,引起了凋落叶中C/N比值的下降。
3 小结与讨论经过2年的模拟氮沉降试验,杉木人工林的年凋落量分别为1 008.83,1 164.10,1 147.30和976.47 kg·hm-2,低氮(N1)和中氮(N2)在一定程度上增加森林凋落物量,而高氮处理(N3)则表现出一定的抑制作用。然而,在模拟氮沉降初期,杉木人工林的年凋落量分别为2 427.51,2 238.10,2 286.66和2 599.50 kg·hm-2,高氮处理(N3)显著增加森林凋落物量(P= 0.034),而中氮(N2)和低氮(N1)则表现出一定的抑制作用(樊后保等,2007)。这2个时期凋落物量对氮沉降增加响应产生差异的原因是,氮沉降一定程度上增加土壤有效氮水平(樊后保等,2007),氮沉降率的增加在一定时间内会提高植物生产力,常见的例子就是林业经营上通过施加氮肥来促进林木生长(Tamm,1991)。但有研究发现,氮输入生态系统的量有一临界值,超出该临界值便会对植物的生产力产生不利影响(Aber et al., 1998),这就造成短期内含氮量高的处理提高森林生产力,但随着时间的推移,当氮沉降超出植物和微生物所需求氮的临界值时,又会表现抑制作用。从本试验的研究结果来看,初步推断120kg N·hm-2 a-1是该杉木人工林的氮饱和点。本研究还发现,在模拟氮沉降试验初期各处理凋落物量要远远大于模拟氮沉降2年后的凋落物量。究其原因,杉木人工林凋落物量可能存在“大小年”现象,3年时间里呈现比较明显的波动性,这与官丽莉等(2004)研究鼎湖山常绿阔叶林凋落物的研究结果相似,这可能是森林维持稳定生产力的一种自我调节形式(张佑倡等,2000)。另外,根据凋落物收集时所记录的天气情况,在2004年试验地所在地区降雨比较多,在5月和7月更是有强台风的影响,这也是造成试验初期凋落物量比较大的主要原因之一。
氮沉降引起凋落物叶C,N元素含量的改变,C /N比值也相应发生了变化,N0处理的C/N比值在1年的变化范围为59.25 ~ 79.29,N1,N2和N3处理分别为57.15 ~ 69.71,50.20 ~ 68.90和43.70 ~61.91,说明经N1,N2和N3处理后,落叶C/N比值的平均值比N0处理分别下降0.27%,5.78%和17.98%。到目前为止,许多学者研究了不同凋落物的性状和可分解性间的关系,发现落叶分解过程中存在氮的绝对积累(Cotrufo et al., 1999),Melin (1930)使用C/N比值来分析落叶的分解特征。C /N比值后来成为评价落叶分解的经典指标(王希华,2004; Sariyildiz et al., 2003)。本研究进行氮沉降对凋落物分解影响的研究发现,除N3处理外,凋落物分解系数与凋落物的C/N比值呈显著的线性关系,表现为C/N比值越低分解越快(樊后保等,2008)。N3处理所表现出来的差异说明氮沉降虽然在一定程度提高凋落物底物的质量,但过高含量的氮沉降水平会引起分解者微生物群落组成向低效率转变,使微生物降解中C的限制加剧等原因,从而延缓凋落物的分解(Micks et al., 2004)。
N0,N1,N2和N3处理凋落物C元素的归还量分别为474.70,544.07,538.55和474.02 kg·hm-2,N元素的归还量分别为7.21,8.56,9.03和9.04kg·hm-2,P元素的归还量分别为1.17,1.24,1.32和1.09 kg·hm-2。森林生态系统中,凋落物是一个重要的碳库,其变化动态将会对全球碳预算产生重大影响。本研究发现,N1,N2处理不仅显著提高凋落物C元素的归还量,同时也提高N,P元素的归还量,N3处理对C,P的归还有一定的抑制作用。这从一方面反映适度的氮沉降可以增加森林生态系统的碳库,高氮处理(N3)则对森林生态系统碳库起抑制作用。然而,有关氮沉降能否增加森林生态系统的碳贮存,在科学界还存在广泛的争议。Townsend等(1996)和Richard等(2004)的研究指出,在北半球温带地区,氮素施肥估计每年增加0.3 ~ 0.5 Pg C贮存。然而,其他的预测结果表明,氮素对生态系统的刺激作用不可能引起明显的碳贮存(Korner,2000),而实际上是减少生态系统生产力和碳贮量(Schulze,1989; Cao et al., 1998)。这些结论的差异和一致性说明氮沉降对森林生态系统碳贮量存在一定的影响,但不同气候类型和树种对氮沉降增加的响应机制可能有所不同。
樊后保, 刘文飞, 李燕燕, 等. 2007. 亚热带杉木(Cunninghamia lanceolata)人工林生长与土壤养分对氮沉降的响应[J]. 生态学报, 27(11): 4630-4641. DOI:10.3321/j.issn:1000-0933.2007.11.031 |
樊后保, 刘文飞, 裘秀群, 等. 2007. 杉木人工林凋落物量对氮沉降增加的初期响应[J]. 生态学杂志, 26(9): 1335-1338. |
樊后保, 刘文飞, 徐雷, 等. 2008. 氮沉降下杉木人工林(Cunninghamia lanceolata)凋落叶分解过程中C、N元素动态变化[J]. 生态学报, 28(6): 2546-2553. |
樊后保, 刘文飞, 杨跃霖, 等. 2008. 杉木人工林凋落物分解对氮沉降增加的响应[J]. 北京林业大学学报, 20(2): 8-13. |
樊后保, 苏兵强, 林德喜, 等. 2000. 杉木人工林生态系统的生物地球化学循环: Ⅱ[J]. 氮素沉降动态.应用与环境生物学报, 6(2): 133-137. |
樊建凌, 胡正义, 庄舜尧, 等. 2007. 林地大气氮沉降的观测研究[J]. 中国环境科学, 27(1): 7-9. |
官丽莉, 周国逸, 张德强, 等. 2004. 鼎湖山南亚热带常绿阔叶林凋落物量20年动态研究[J]. 植物生态学报, 28(4): 445-456. |
黄忠良, 丁明懋, 张祝平, 等. 1994. 鼎湖山季风常绿阔叶林的水文学过程及其氮素动态[J]. 植物生态学报, 18(2): 194-199. |
王希华, 黄建军, 闫思荣. 2004. 天童国家森林公园常见植物凋落叶分解的研究[J]. 植物生态学报, 28(4): 457-467. DOI:10.17521/cjpe.2004.0063 |
肖健. 2005. 漳州市氮湿沉降量异常的形成及危害[J]. 能源与环境, (2): 59-61. |
张佑倡, 孔国辉, 余清发, 等. 2000. 鼎湖山演替系列中代表性森林凋落物研究[J]. 生态学报, 20(6): 938-944. |
周国逸, 闫俊华. 2001. 鼎湖区域大气降水特征和物质元素输人对森林生态系统存在和发育的影响[J]. 生态学报, 21(12): 2002. DOI:10.3321/j.issn:1000-0933.2001.12.006 |
Aber J D, McDowell W, Nadelhoffer K J, et al. 1998. Nitrogen saturation in northern forest ecosystems[J]. Bioscience, 39(6): 378-386. |
Cao M, Woodward F I. 1998. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change[J]. Nature, 393: 249-252. DOI:10.1038/30460 |
Christopher S. 1999. Terrestrial biomass and the effect of deforestation on the globe carbon cycle[J]. Bioscience, 49: 769-778. DOI:10.2307/1313568 |
Cotrufo M F, Rasehi A, LanIni M, et al. 1999. Decomposition and nutrient dynamics of Quercus pubescens leaf litter in a naturally enriched CO2 Mediterranean ecosystem[J]. Functional Ecology, 13(3): 343-351. DOI:10.1046/j.1365-2435.1999.00328.x |
Fang H, Mo J M. 2006. Effect of nitrogen deposition on forest litter decomposition[J]. Acta Ecologica Sinica, 26(9): 3127-3136. |
Galloway J N, Schlesinger W H, Levy I I, et al. 1995. Nitrogen fixation: anthropogenic enhancement-environmental response[J]. Global Biogeochemical Cycles, 9(2): 235-252. DOI:10.1029/95GB00158 |
Galloway J N, Dentener F J, Capone D G, et al. 2004. Nitrogen cycles: past, present, and future[J]. Biogeochemistry, 70(2): 153-226. DOI:10.1007/s10533-004-0370-0 |
Kochy M, Wilson S D. 2001. Nitrogen deposition and forest expansion in the northern great plains[J]. Journal of Ecology, 89(5): 807-817. DOI:10.1046/j.0022-0477.2001.00600.x |
Korner C. 2000. Biosphere responses to CO2 enrichment[J]. Ecology Application, 10(6): 1590-1619. |
Melin E. 1930. Biological decomposition of some types of litter from North American forests[J]. Ecology, 11(1): 72-101. DOI:10.2307/1930782 |
Micks P, Downs M R, Magill A H. 2004. Decomposition litter as a sink for 15N-enriched additions to an oak forest and a red pine plantation[J]. Forest Ecosystem and Management, 196(1): 71-87. DOI:10.1016/j.foreco.2004.03.013 |
Nihlgard B. 1985. The ammonium hypothesis: an additional explanation to the forest die back in Europe[J]. Ambio, 14: 2-8. |
Palviainen M, Finér L, Kurka A M, et al. 2004. Release of potassium, calcium, iron and aluminium from Norway spruce, Scots pine and silver birch logging residues[J]. Plant and Soil, 259(1/2): 123-136. DOI:10.1023/B:PLSO.0000020938.78215.bd |
Prescott C E, Blevins L L. 2004. Litter decomposition in British Columbia forests influences of forestry activities[J]. Journal of Ecosystems and Management, 5(2): 30-43. |
Richard D, Bowdena E D, Kathleen S, et al. 2004. Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard forest[J]. Forest Ecology and Management, 196(1): 43-56. DOI:10.1016/j.foreco.2004.03.011 |
Sariyildiz T, Anderson J M. 2003. Interactions between litter quality, decomposition and soil fertility: a laboratory study[J]. Soil Biology and Biochemistry, 35(3): 391-399. DOI:10.1016/S0038-0717(02)00290-0 |
Schulze E D. 1989. Air pollution and forest decline in a spruce(Picea abies) forest[J]. Science, 244: 776-783. DOI:10.1126/science.244.4906.776 |
Tamm C O. 1991. Nitrogen in terrestrial ecosystems: Questions of productivity, vegetational changes, and ecosystem stability. Berlin: Springer-Verlag, 50-97.
|
Townsend A R, Braswell B H, Holland E A, et al. 1996. Spatial and temporal patterns in terrestrial carbons storage due to deposition of fossil fuel nitrogen[J]. Ecological Applications, 6(3): 806-814. DOI:10.2307/2269486 |
Vejre H, Callesen I, Vesterdal L, et al. 2003. Carbon and nitrogen in Danish forest soils-contents and distribution determined by soil order[J]. Soil Science Society of America Journal, 67(1): 335-343. DOI:10.2136/sssaj2003.3350 |
Vitousek P M, Mooney H A, Lubchenco J, et al. 1997. Human domination of earth's ecosystems[J]. Science, 277: 494-499. DOI:10.1126/science.277.5325.494 |