 2011, Vol. 47
2011, Vol. 47文章信息
- 张艳杰, 温佐吾
- Zhang Yanjie, Wen Zuowu
- 不同造林密度马尾松人工林的根系生物量
- Root Biomass of Pinus massoniana Plantations under Different Planting Densities
- 林业科学, 2011, 47(3): 75-81.
- Scientia Silvae Sinicae, 2011, 47(3): 75-81.
-
文章历史
- 收稿日期:2009-08-13
- 修回日期:2010-12-05
-
作者相关文章



2. 贵州大学林学院 贵阳 550025
2. Forestry College, Guizhou University Guiyang 550025
根系是树木最重要营养器官,不但为林木地上部分提供固定和机械支撑作用,从土壤中吸收养分和水分,而且通过呼吸和周转,消耗光合产物并向土壤输入有机质,对林木生长起着决定性作用(Canndeli et al., 1996; 王政权等,2008),在发挥林木功能和森林生态系统能量流动和物质循环中扮演重要角色。从20世纪中期以来,随着人类对生物地球化学循环过程的重视,森林根系的研究才受到关注,它的研究加深人们对森林生态系统功能和效益的了解(贺金生等,2004)。国外许多学者十分重视对林木根系生长及其生物量的研究(伯姆,1985; Casper et al., 1997; Burkes et al., 2003),我国一些学者也对华北落叶松(Larix principis-rupprechtii)、赤松(Pinus densiflora)、油松(Pinus tabulaeformis)、小叶杨(Populus simonii)、檫木(Sassafras tzumu)、云南松(Pinus yunnanensis)、水曲柳(Fraxinus mandschurica)等树种的根系生物量进行了研究(韩有志等,1997; 杜晓军等,1998; 王成等,1999; 梁建萍等,1999; 崔浪军等,2003; 朱慧等,2003; 邓坤枚等,2005; 梅莉等,2006)。
马尾松(Pinus massoniana)是我国南方广泛分布的用材林树种,在森林资源构成中占有十分重要的地位。近些年来,一些学者对马尾松人工林、飞播林和天然次生林生物量的结构特征、分配规律及生产力水平等进行了研究,其中也包括对林木和林分根系生物量的测定与研究(冯宗炜等,1982; 田大伦等,1986; 刘煊章,1993; 董志彬,2000; 陈兆先等,2001; 丁贵杰,2003; 陈美高,2006; 张治军等,2006; 2008; 漆良华等,2007),但对马尾松林分根系生物量进行专门研究的报道却很少,对不同造林密度且林龄较大的马尾松林分进行根系生物量专门研究的报道更少。
本文以贵州省龙里林场20年生马尾松造林密度试验林为研究对象,采用全根挖掘法和土钻法(伯姆,1985; 黄建辉等,1999),对5种栽植密度试验样地的平均标准木单木根系生物量和林分行间根系生物量分别进行了测定和研究,为了解不同密度马尾松人工林根系生物量及其空间分布规律,提高马尾松人工林的经营管理水平提供科学依据。
1 试验地概况试验地位于贵州省龙里林场哨上工区,属马尾松分布的中带西区。气候属中亚热带温和湿润类型,年平均气温14.8 ℃,年平均降水量1 089.3 mm,年平均相对湿度79%。试验地海拔约为1 150 m,地貌为侵蚀、剥蚀低中山台地,坡向东南,坡度5 ~10°。母岩为石英砂岩,土壤为薄腐殖质层轻质厚层山地黄壤。马尾松立地指数16左右,立地质量中等。试验地前作为马尾松人工林,林下灌木、草本主要为茅栗(Castanea seguinii)、小叶南烛(Vaccinium bracteatum var.chinense)、滇白珠(Gaultheria leucocarpa var.crenulata)、胡枝子(Lespedeza bicolor)、蕨(Pteridium aquilinum var.latiusculum)、五节芒(Miscanthus floridulus)、铁芒萁(Dicranopteris linearis)等。
2 研究方法 2.1 试验设计造林密度试验采用随机区组设计,设置5个处理,每个处理3次重复,共15个固定样地,试验小区面积为400 m2 (20 m × 20 m)。设计的5个处理株行距分别为3 m × 3 m (1 111株·hm-2),2 m × 2 m (2 500株·hm-2),1.5 m × 1.5 m (4 444株·hm-2),1 m × 1 m (10 000株·hm-2),0.7 m × 0.7 m (20 408株·hm-2),分别用A,B,C,D,E来表示。
造林施工采用全面整地、拉线定点、逐点挖穴定植的方法。栽植穴的规格为40 cm × 40 cm × 25 cm。供试苗木为当地种源,自育1年生1,2级苗木造林。苗木平均高24.1 cm,平均地径为0.25 cm。造林时间为1983年2月,经成活率调查后,对少数死亡的苗木,于翌年春用同龄苗木进行补植。造林当年砍草抚育2次,第2,3年分别进行松土除草各2次。在整个试验过程中,林分均未进行过抚育间伐,但由于林分的自然稀疏和某些人为破坏,在林龄20年调查时,A,B,C,D,E各处理林分的平均保存密度分别降低为908,1 830,3 585,5 750,7 436株·hm-2。
2.2 根系生物量研究方法本次研究在参考伯姆(1985)、王成等(1999)分类办法的基础上,结合在根系挖掘和调查测定中的实际情况,将不同根系分为细根(根径<0.2 cm)、小根(根径0.2 ~ 0.5 cm)、中根(根径0.5 ~ 1.0cm)、大根(根径1.0 ~ 2.0 cm)、粗根(根径>2.0cm)和根桩6类。
采用全根挖掘法测定林分平均标准木单木生物量,每个样地的林分平均标准木均从离地表 2 cm左右处伐倒,在完成地上部分生物量测定后,用锄头尽可能挖掘出全部根桩、粗根、大根、中根及小根,将其仔细清理干净后,称量各级根的鲜质量,然后分别采集50 ~ 60 g样品用于测定水分含量。
|
|
采用土钻法测定林分行间生物量,土钻为一内径5.04 cm、外径5.71 cm、长度60 cm、头部带锯齿的无缝钢管。在每个样地内的林木行间随机设置20个采样点,每个采样点每隔10 cm为一个层次钻取土样,一直钻到50 cm深度。从采集到的土样中分层分离出细根、小根和部分中根3个不同径级根系,分别清理干净后称其鲜质量,并将其作为测定含水量的样品。
所有样品带回实验室后,在105 ℃恒温下烘干至恒量,测定各样品的含水率,推算出各组分的生物量。
2.3 数据处理对不同密度林分平均标准木单木及林分行间不同级别的根系生物量数据,均运用SPSS软件进行方差分析处理。对方差分析结果差异显著或接近显著者,则采用LSR检验法进行各平均值两两间的多重比较。此外,对不同密度林分间不同级别根系生物量的平均值,还分别计算其标准差(陈华豪等,1988)。
3 结果与分析 3.1 林分平均标准木单木根系生物量从表 1可以看出,5种不同的造林密度,其林分平均标准木单木的根系生物量,均以根桩所占比例最大,占根系总生物量的58.30% ~ 72.46%,其中密度最小的A处理所占比例最低。粗根生物量占根系总生物量的18.18% ~ 37.87%,其中密度最小的A处理,其粗根生物量占根系总生物量的比例为密度最大的E处理的2.08倍。根桩和粗根两者的生物量是平均标准木单木根系总生物量的主体,其两者之和占根系总生物量的比例,A,B,C,D,E 5种密度分别为96.17%,96.34%,92.77%,94.01%和90.64%,这表明马尾松林木在生长过程中,需利用根桩和粗根发挥支撑和固定树体的作用。除根桩和粗根外,大根、中根和小根生物量占根系总生物量的比例则较低。不同密度处理林分平均标准木单木的大根生物量为2.32% ~ 4.96%,中根生物量为0.74% ~ 3.25%,小根生物量为0.09% ~ 2.25%。5种不同造林密度的马尾松林分,其不同级别根系生物量的排序基本上都为:根桩>粗根>大根>中根>小根。
|
|
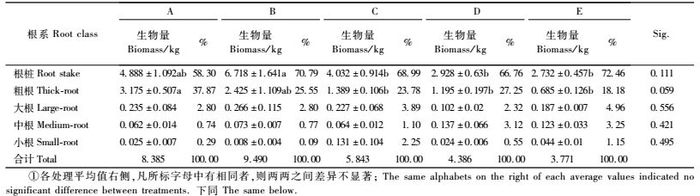
表 1中方差分析和多重比较的结果表明,在林分平均标准木单木的各级根系中,根桩生物量在不同密度林分之间仅在接近0.1的水平上有差异,其中只有B密度林分根桩生物量与其余4种密度之间的差异为显著。粗根生物量在不同密度林分之间的差异,接近于达到显著水平,其中: A,B 2个密度林分生物量之间的差异不显著,但A密度林分与C,D,E 3种密度粗根生物量之间的差异达到显著水平。而对于大、中、小根的生物量,林分的不同密度对其则无显著影响。
由表 1可知,在马尾松不同造林密度情况下,平均标准木单木各级根系生物量随密度的增加而出现不同的变化。除密度最小的A处理之外,随着林分密度的增加,B,C,D,E 4种密度处理林分平均标准木单木的总生物量大体上呈现逐渐减少的规律,这表明对根系的生长及其生物量的积累而言,林分的密度效应比较明显。粗根生物量均随林分密度的增大而减小,最小密度A处理的粗根生物量约为最大密度E处理的4.6倍。大根生物量大体上随林分密度的增大先增加、后减小,在D密度时降到最小,到E密度时又再度增加,其最大生物量为B处理(0.266 kg),最小为D处理(0.102 kg)。中根生物量的变化较为平缓,最大值出现在密度较高的D处理(0.137 kg),最小值为密度最小的A处理(0.062kg)。小根生物量的峰值出现在中等密度的C处理(0.131 kg),其余4种处理的小根生物量较小,在0.008 ~ 0.044 kg。
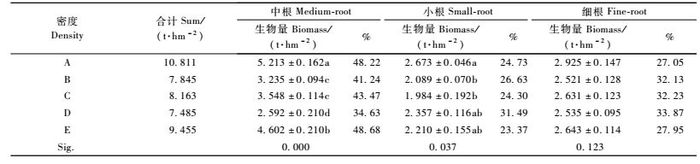
3.2 林分行间根系生物量由表 2可知,A,B,C,D,E 5种不同的造林密度,其林分行间根系生物量中的中根、小根及细根的生物量,均未随林分密度的增加而出现规律性的变化。生物量由大到小的排序,中根是A>E>C>B> D,其中B,C 2种密度相差不大,小根是A>D>E> B>C,细根则是A>E>C>D>B。此外,生物量最高和最低的2个密度林分之间的差异,中根生物量为101.1%,小根生物量是34.7%,细根生物量则仅为16.0%,也就是说,行间根系生物量在不同密度林分之间的差异,从中根到小根再到细根明显减小,这说明林分的不同密度对中根、小根和细根生物量生长的影响,出现逐渐变小的趋势。无论是何种造林密度,林分行间不同级别根系的生物量均存在中根>细根>小根的规律性。中根、细根和小根在合计生物量中所占的比例,分别是34.63% ~48.68%,27.05% ~ 33.87%和23.37% ~ 31.49%。
从表 2中方差分析的结果可以看出,在林分行间的3个级别根系中,中根生物量在不同密度林分之间的差异达到极显著水平,小根生物量达到显著水平,而细根生物量则未达到显著水平。多重比较的结果表明,5种不同密度林分的中根生物量,除B,C 2种密度林分两两之间的差异未达到显著水平外,其余A密度林分与E,B,C,D 4种密度林分之间,E密度林分与B,C,D 3个密度林分之间,B,C密度林分与D密度林分之间,其生物量的差异均达到显著水平。5种不同密度林分的小根生物量,只有A密度林分与B,C,D,E 4种密度林分之间的差异达到显著水平,而其余4种密度林分相互之间的差异则均未达到显著水平。
3.3 林分行间根系生物量随土层深度的分布规律从表 3中林分行间不同级别根系生物量在不同土层深度的垂直分布情况可以看出,按照A,B,C,D,E 5种处理林分密度从小到大的顺序,0 ~ 10 cm土层根系生物量分别占各层根系总生物量的35.64%,46.17%,27.77%,38.89%和38.12%,10 ~ 20 cm土层分别占38.97%,26.63%,33.16%,29.89%和28.73%,20 ~ 30 cm土层分别占18.14%,17.99%,21.48%,18.28%和18.44%。0 ~ 30 cm土层根系生物量之和,则分别占5个土层根系总生物量的92.75%,90.79%,82.41%,87.06%和85.29%。由此可见,20年生不同造林密度马尾松人工林的根系,主要集中分布在0 ~ 30 cm的表层土壤中,占根系总生物量的82%以上,而30 ~ 50 cm土层的根系生物量所占比例很小,不足18%。从表 3还可知,随着土层深度的增加,各土层行间根系总生物量呈逐渐减少的趋势。在0 ~ 30 cm的土层中,密度最小的A密度林分根系生物量最大,密度最大的E密度林分次之,这主要是由于密度小的林分各单株根系有充足的生长空间,而密度最大林分因其林木株数最多从而使其根系生物量较大。
|
|
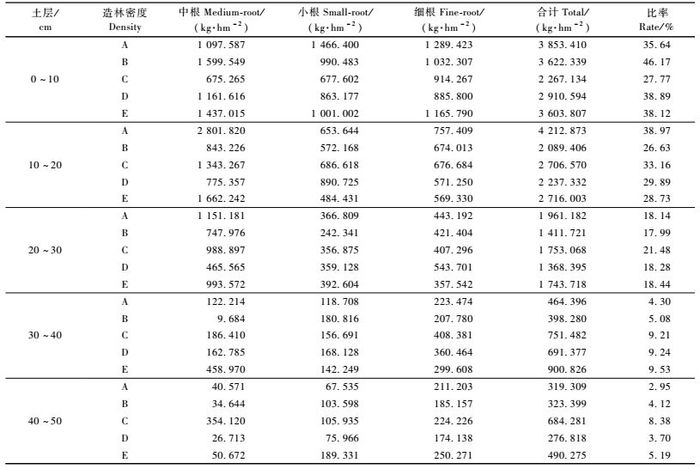
从表 3中还可以看出,A,B,C,D,E 5种密度林分根系行间生物量由大到小的排序,在0 ~ 10 cm的土层,中根是B>E>D>A>C,小根是A>E>B >D>C,细根是A>E>B>C>D; 在10 ~ 20 cm的土层,中根是A>E>C>B>D,小根是D>C>A>B> E,细根是A>C>B>D>E; 在20 ~ 30 cm的土层,中根是A>E>C>B>D,小根是E>A>D>C> B,细根是D>A>B>C>E; 在30 ~ 40 cm的土层,中根是E>C>D>A>B,小根是B>D>C>E> A,细根是C>D>E>A>B; 在40 ~ 50 cm的土层,中根是C>E>A>B>D,小根是E>C>B>D> A,细根是E>C>A>B>D。在同一土层内,5个不同的造林密度,其林分行间根系的生物量,未出现随林分密度增减的明显规律性变化。
从表 3可进一步得知,5种不同造林密度的林分,其林分行间不同级别根系生物量随土层加深而产生的变化趋势各自存在一定的规律性。密度最大和最小的A,E 2个密度林分,除0 ~ 10 cm土层中根生物量小于10 ~ 20 cm土层外,其余各土层的中根和所有各土层的小根和细根的生物量,大体上均随土层的加深而减少。B,D 2个密度林分,其中根、小根和细根的生物量也大体上随土层的加深而减少。C密度林分的中根生物量在10 ~ 20 cm土层中最大,20 ~ 30 cm土层次之,30 ~ 40 cm土层最少,小根和细根的生物量则大体上随土层的加深而减少。
3.4 根系生物量与林木胸径、树高及林分密度的相关关系以不同径级的根系生物量(W)为因变量,林木胸径(D)、树高(H)及林分密度(De)为自变量,采用多元回归模型拟合不同造林密度马尾松林分平均标准木单木各级根系生物量与胸径、树高及密度关系的数学模型(陈兆先等,2001; 朱慧等,2003),拟合的结果见表 4。从表 4可知,林分平均标准木单木各级根系生物量与林木胸径、树高及林分密度的相关关系较为密切,所拟合的各个回归模型的相关系数除小根略小于0.8外,其余均在0.8以上,精度比较高。
|
|

再以各级根系生物量(W)为因变量,林分密度(x)为自变量,采用指数函数模型拟合不同造林密度马尾松林分行间各级根系生物量与林分密度关系的数学模型,拟合的结果见表 5。从表 5可知,林分行间各级根系生物量与林分密度的相关关系也比较密切,所拟合的3个回归模型的相关系数除中根较低外,其余均在0.90以上,说明林分行间根系的生物量受林分密度的影响较大。
|
|

1) 林分平均标准木单木不同级别根系的生物量,5种不同造林密度马尾松林分的排序为:根桩>粗根>大根>中根>小根。其中根桩和粗根是根系总生物量的主体,两者之和占根系总生物量的比例较大,5种密度林分均达90%以上。根系总生物量在2 500 ~ 20 408株·hm-2的范围内随林分密度的增高逐渐减少,这表明对根系的生长及其生物量的积累而言,林分的密度效应比较明显。
2) 5种不同造林密度马尾松林分行间的中根、小根及细根的生物量,未出现随林分密度的增加而发生规律性变化的现象。但林分的不同密度对中根、小根和细根生物量生长的影响,则出现逐渐变小的趋势。此外,5种不同密度的林分,其行间不同级别根系的生物量均存在中根>细根>小根的规律性。细根是各级根系中最小的根系,但其生物量在林分行间根系生物量中所占的比例却并不是最低,达到27.05% ~ 33.87%,这表明其在外生菌根的帮助下从土壤中吸收水分和养分,参与物质循环,以提高林分的生产力(崔浪军等,2003)。
3) 一般来说,林木根系随着土壤层次的加深而大幅度减少(Leuschner et al., 2004; Schenk et al., 2002)。本研究中5种不同密度马尾松林分的根系主要分布在0 ~ 30 cm的表层土壤中,占根系总生物量的82%以上。而随着土层深度的增加,不同密度林分行间根系生物量也出现逐渐减少的趋势。但在同一土层内,其根系生物量则未出现随林分密度增减明显规律性变化。
4) 不同数学模型拟合的结果表明,林分平均标准木单木各级根系生物量与林木胸径、树高及林分密度的相关关系较为密切,林分行间各级根系生物量与林分密度成指数函数关系,其相关关系也比较密切,说明林分根系生物量生长受林木胸径、树高和林分密度的影响较大。
5) 如前所述,对马尾松林木和林分的根系生物量,已有一些学者进行过研究。但这些研究多是在现有的马尾松人工林、飞播林或天然次生林中设置临时调查样地,其研究的内容虽然都涉及到不同径级、不同层次、不同密度、不同年龄、不同优势度、不同立地的林木或林分的根系生物量,但这些研究却多是以地上部分生物量为主,根系生物量为辅,很少有专门针对根系生物量进行的研究,更无在试验设计基础上对固定样地马尾松林分所进行的根系生物量研究。本文的研究以设置、观测20年生马尾松造林密度试验林固定样地为基础,对马尾松林木的根系生物量进行专门的调查研究,研究内容既包括各样地不同密度林分平均标准木单木的根系生物量,也包括林分行间的根系生物量。既研究不同密度林分平均标准木单木不同级别根系生物量的分配规律,又研究不同密度林分行间根系生物量在不同土壤层次中的空间分布特征。由于试验样地设置5种不同的密度处理,每个处理又设置3个重复,因此,本文还对样地不同处理林分平均标准木单木的根系生物量和林木行间的根系生物量测定数据进行包括平均值标准差、方差分析和多重比较在内的统计分析。此外,还通过回归分析,初步探讨根系生物量与林分密度之间的相关关系。这些研究内容,是其他学者在马尾松根系生物量研究中很少或没有涉及到的。
6) 本研究是在5种不同造林密度试验林内进行,由于马尾松是深根性树种,且在不同密度样地上其根系的生长盘根错节,在采用全根挖掘法挖取林分平均标准木单木根系时,难于做到将最小的细根百分之百地彻底挖净,这就使本项研究的结果可能产生某些误差,需在以后的研究中加以完善。
伯姆W. 1985. 根系研究法. 薛德榕, 谭协麟译. 北京: 科学出版社.
|
陈华豪, 丁恩统, 蔡贤如, 等. 1988. 林业应用数理统计[M]. 大连: 大连海运学院出版社.
|
陈美高. 2006. 不同年龄马尾松人工林生物量结构特征[J]. 福建林学院学报, 26(4): 332-335. |
陈兆先, 何友军, 柏方敏, 等. 2001. 林分密度对马尾松飞播林生物产量及生产力的影响[J]. 中南林学院学报, 21(1): 44-47. |
崔浪军, 梁宗锁, 韩蕊莲, 等. 2003. 沙棘-杨树混交林生物量、林地土壤特性及其根系分布特征研究[J]. 林业科学, 39(6): 1-7. DOI:10.11707/j.1001-7488.20030601 |
邓坤枚, 罗天祥, 张林, 等. 2005. 云南松林的根系生物量及其分布规律的研究[J]. 应用生态学报, 16(1): 21-24. |
丁贵杰. 2003. 马尾松人工林生物量和生产力研究[J]. 福建林学院学报, 23(1): 34-38. |
董志彬. 2000. 马尾松、杜仲混交林的根系与生物量研究[J]. 滁州师专学报, 2(4): 95-97. |
杜晓军, 刘常富, 金罡, 等. 1998. 长白山主要森林生态系统根系生物量研究[J]. 沈阳农业大学学报, 29(3): 229-232. |
冯宗炜, 陈楚莹, 张家武, 等. 1982. 湖南会同地区马尾松林生物量的测定[J]. 林业科学, 18(2): 127-134. |
韩有志, 姚延, 梁建萍, 等. 1997. 华北落叶松人工林根系生物量的研究[J]. 林业科技通讯, (7): 17-19. |
贺金生, 王政权, 方精云. 2004. 全球变化下的地下生态学:问题与展望[J]. 科学通报, 49(13): 1226-1233. DOI:10.3321/j.issn:0023-074X.2004.13.002 |
黄建辉, 韩兴国, 陈灵芝. 1999. 森林生态系统根系生物量研究进展[J]. 生态学报, 19(2): 270-277. |
梁建萍, 韩有志, 张云龙, 等. 1999. 油松人工林根系生物量的研究[J]. 河南科学, 17(增l): 77-79. |
刘煊章. 1993. 不同年龄马尾松林生物量的研究[J]. 林业资源管理, (2): 77-80. |
梅莉, 王政权, 韩有志, 等. 2006. 水曲柳根系生物量、比根长和根长密度的分布格局[J]. 应用生态学报, 17(1): 1-4. |
漆良华, 张旭东, 周金星, 等. 2007. 马尾松飞播林生物量与生产力的变化规律与结构特征[J]. 林业科学研究, 20(3): 344-349. |
田大伦, 潘维俦. 1986. 马尾松林杆材阶段生物产量和径级分化及密度效应初探[J]. 植物生态学与地植物学学报, 10(4): 294-301. |
王成, 金永焕, 刘继生, 等. 1999. 延边地区天然赤松林单木根系生物量的研究[J]. 北京林业大学学报, 21(1): 44-49. |
王政权, 郭大立. 2008. 根系生态学[J]. 植物生态学报, 32(6): 1213-1216. |
张治军, 王彦辉, 于澎涛, 等. 2008. 不同优势度马尾松的生物量及根系分布特征[J]. 南京林业大学学报, 32(4): 71-75. |
张治军, 王彦辉, 袁玉欣, 等. 2006. 马尾松天然次生林生物量的结构与分布[J]. 河北农业大学学报, 29(5): 37-43. |
朱慧, 洪伟, 吴承祯, 等. 2003. 天然更新的檫木林根系生物量的研究[J]. 植物资源与环境学报, 12(3): 31-35. |
Canndeli J, Jackson R B, Ehleringer J R, et al. 1996. Maximum rooting depth of vegetation types at the global scale[J]. Oecllogia, 108(4): 583-595. DOI:10.1007/BF00329030 |
Casper B B, Jackson R B. 1997. Plant competion underground[J]. Annual Review of Ecology and Systematics, 28: 545-570. DOI:10.1146/annurev.ecolsys.28.1.545 |
Burkes E C, Will R E, Barron-Gafford G A, et al. 2003. Biomass partitioning and growth efficiency of intensively managed Pinus Taeda and Pinus elliottii stands of different planting densities[J]. Forest Science, 49(2): 224-234. |
Leuschner C, Hertel D, Schmid I, et al. 2004. Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility[J]. Plant and Soil, 258(1/2): 43-56. |
Schenk H J, Jackson R B. 2002. The global biogeography of roots[J]. Ecol Mono, 72(3): 311-328. DOI:10.1890/0012-9615(2002)072[0311:TGBOR]2.0.CO;2 |