 2011, Vol. 47
2011, Vol. 47文章信息
- 亓玉飞, 尹伟伦, 夏新莉, 孙尚伟
- Qi Yufei, Yin Weilun, Xia Xinli, Sun Shangwei
- 修枝对欧美杨107杨水分生理的影响
- Effects of Pruning on Water Physiology of Poplar Clone Populus × euramericana cv. '74 /76'
- 林业科学, 2011, 47(3): 33-38.
- Scientia Silvae Sinicae, 2011, 47(3): 33-38.
-
文章历史
- 收稿日期:2010-01-23
- 修回日期:2010-06-24
-
作者相关文章



2. 北京林业大学教育部森林培育与保护重点实验室 北京 100083;
3. 中国林业科学研究院 北京 100091
2. Key Laboratory for Silviculture and Conservation, Ministry of Education Beijing Forestry University Beijing 100083;
3. Chinese Academy of Forestry Beijing 100091
我国杨树(Populus)人工林面积约667万hm2,占全国人工林面积的20%,并且70%以上的杨树人工林为黑杨派树种(张津林等,2006)。欧美杨107杨(Populus×euramericana cv.‘74 /76’)作为我国重要的黑杨派速生林树种,目前已经在河北、山东、河南等多省进行大面积的推广和栽植。已有研究表明:作为林木抚育的基础性措施,合理修枝在改善树木干形使其更加圆满,提高造林成活率、保存率,提高林地生产力,增加材积生长量和木材利用率等方面具有重要作用(陈森琨等,2008; 孙尚伟等,2008)。根据杨树无性系的生长特性、林分密度、林分年龄以及立地条件(特别是水分和养分条件)如何来确定适宜的修枝强度是生产中急需解决的问题。本文通过对不同年龄和不同修枝强度处理下,欧美107杨的树干液流(stem sap flow)、蒸腾速率(Tr)及叶水势(LWP)等水分生理变化的研究,并结合不同修枝强度对107杨生长的影响(陈森琨等,2008),为107杨在不同生长阶段确定合理的修枝强度提供支撑。
1 试验地自然概况试验地位于河北省保定市望都县赵庄乡速生丰产林基地,地处河北省中西部,位于38° 30′46″— 38°48′30″ N,115°01′16″—115°18′13″ E之间。属于北温带半干旱季风气候; 年平均日照2 580 h; 年平均气温12.3 ℃,7月平均气温26.5 ℃,1月平均气温-4.1 ℃; 年平均降水量473 mm; 年均无霜期191天。土壤主要为潮褐土。
2 材料与方法 2.1 试验材料试验地林木为欧美杨107杨,树行南北走向。试验地Ⅰ:2006年定植,株行距为3 m×4 m,为4年样木; 试验地Ⅱ:2004年定植,株行距为4 m×2 m,为6年样木。
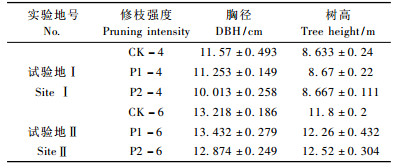
试验地Ⅰ,Ⅱ都采取完全随机区组试验设计。通过每木检尺,选取径阶平均木为样木,要求干形通直圆满、不偏心、树冠良好且不偏冠。2块试验地都设置3个处理组,每个处理组都设有3种处理,即对照处理CK(不修枝)、中度处理P1(修掉树冠高度的1 /3)、重度修枝P2(修掉树冠高度的2 /3)。2块试验地都设有保护行,以提高试验精确度。2008年春末对试验样木进行修枝处理,2009年春末进行本试验的各项指标的测定。试验样木的树高、胸径形态指标见表 1。
|
|
采用美国进口Licor-6400光合便携仪测定蒸腾速率(Tr)生理指标。在生长季内,每个月选择晴好天气2天,在9:00—11:00间选取处理样木上部枝条,测定从枝条顶端起第3,5,7,10叶片,每次测量重复3次,结果取平均值。4年生107杨使用脚手架辅助工具,原位测定各项指标; 6年生107杨,由于树木高大,采用取样枝的方法测定各项指标。利用CO2钢瓶提供稳定ϕ(CO2)=400×10-6气体,利用licor 6400-02B人工光源,设定光合有效辐射(PAR)为1 500 (μmol·m-2s-1)。
叶水势(LWP)由英国进口的Skye植物水势压力室仪测定。提供压力气体为氮气(N2),压力单位Bar(1 Mpa=10Bar)。叶水势在测蒸腾速率当天清晨6:00—7:00之间测定。选取各处理样木上部枝条,取从枝条顶端起第3,5,7叶片进行测定,结果取平均值。
树干液流采用Greenspan茎流计进行测定。在生长季内持续不间断测定记录液流通量及液流速率。Greenspan茎流计是以热脉冲技术和热补偿理论为原理进行设计的目前国内外较为精确的树干液流测定仪(王华田等,2002b)。探针测定部位位于树干南向胸径1.3 m处。安装探头(SF100)时,将钻孔模块(有3个直径为3 mm的小孔,上孔与中孔相距10 mm,中孔与下孔相距为5 mm)固定在待测点上,用微电钻机沿模块打孔,深度为上、下孔2 cm,中孔3 cm。树干边材面积用生长锥以及树干圆盘截取法测定,边材面积为65.97 cm2 (4年)、98.8 cm2 (6年)。试验测定时间设定为每10 min记录数据1次,脉冲持续时间为1.6 s,试验期间24 h连续监测记录。液流通量(Q)为单位时间内通过截面积的液流体积,是液流速率(Vs)与通过液流的树干边材面积(A)的乘积。Q=VsA。
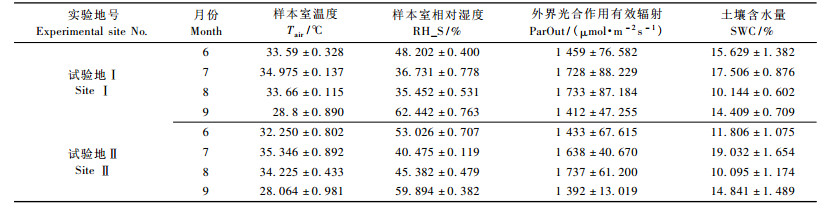
生理指标测定当天的气候因子由licor-6400光合便携仪测定提供。本试验仪器所测指标:样本室温度Tair(℃),样本室相对湿度RH_S(%),外界光合作用有效辐射ParOut(μmol·m-2s-1),代表着水分生理指标测定当天的气候因子状况。
采用土钻烘干法测定试验地0~40 cm范围内的土壤含水量,每10 cm为一层,分4层。测定点设在距样木南向30 cm处。每层按3种处理重复3次,取其均值。每个月水分生理指标测定当天进行测定。
3 结果与分析 3.1 生理指标测定当天气候因子及土壤含水量值林木水分生理指标变化与气候因子(太阳辐射强度、空气温度和空气相对湿度等)密切相关(王华田等,2002a),与土壤水分状况也有一定关系(Gullo et al., 1992)。结合表 2分析:1) 6—8月,样本室温度Tair较高,样本室相对湿度RH_S较低,外界光合作用有效辐射较高。根据兰州干旱气象研究所干旱监测预测研究室报告可知:试验期间温度比同期偏高1 ℃以上。由河北省石家庄水文局报告可知:平均降水量376 mm,较08年同期减少53.7%。综合分析,生长季(6—8月)内,整个试验地处于干旱状态。2)9月Tair降低,RH_S变高,ParOut变低; 由水文局报告可知:平均降水量与常年同期偏多30%,与往年相比,试验地保持正常状态。3)试验地Ⅰ,Ⅱ土壤含水量在生长季内都处在20%以下,随着土壤深度的增加,土壤含水量基本不变。相较往年土壤含水量均值来说,土壤含水量低,属于缺水年份。试验期间气候因子及土壤含水量值见表 2。
|
|
树干液流传导是研究树木水分变化的重要指标。从图 1和图 2可知:生长季(6—9月)内,修枝处理和未修枝对照样木树干液流连日变化有着明显的规律性。1) 3种处理液流日变化基本皆呈现单宽峰曲线,其波动特征基本符合其他研究试验得出的液流日变化变化规律(马履一等,2002)。2) 3种处理的液流通量及液流速率波峰和波谷出现的时间基本相同,而波峰峰值却相差较大。就日变化波峰而言,6,7和8月皆是呈现P1 > CK > P2,9月则呈现CK > P1 > P2的明显规律,而波谷则皆呈现低水平,变化基本一致。3)综合分析可知:在干旱状态(6—8月)时,CK(对照未修枝)液流速率及液流通量处于较低水平; 在正常状态(9月),CK则升至最高水平。这说明干旱使CK呈现缺水状态,不能满足林木的正常生命活动需要,导致树木的生理机能一定程度的减退。因此,CK由于本身吸水等生理能力的下降,致使树干液流速率及液流通量大大降低。当树木恢复正常不缺水状态,树干液流也恢复到最高水平。4)无论在干旱或正常状态,P1 (修枝1 /3)都保持较一致的且较高的液流速率和液流通量。P1修枝处理下,树木即使在环境干旱情况下仍保持着正常的生理机能,进行正常吸水等生命活动。陈森琨等(2008)的研究结果表明:中度修枝P1可以让欧美杨107杨木材生长量显著增加。因此,初步认为中度修枝P1 (修枝1/3),使树木干旱耐受性强,并且生长良好,是较为合适的修枝强度。5) P2 (修枝2/3)在干旱或正常状态下始终处于很低的水平,重度修枝P2使杨树材积生长量大为降低。这说明重度修枝P2(修枝2/3),可能是因为修枝过度枝叶损失过多,导致水分及营养等供应不足,严重影响树木的生长。

|
图 1 生长季内6 a生杨液流通量及液流速率日变化对比 Figure 1 The comparison of Flux and velocity of 6-year-old poplar during growing season A: 6月June; B: 7月July; C: 8月Augest; D: 9月September下同。The same below. |

|
图 2 生长季内4年生杨液流通量及液流速率日变化对比 Figure 2 The comparison of Flux and velocity of 4-year-old poplar during the growing season |
蒸腾强度影响着植物水分状况,蒸腾速率(Tr)的高低,反映了树木的水分生理状况以及植物适应环境能力的大小(曾凡江等,2002)。从图 3可知:生长季内试验样木的3种处理的蒸腾速率(Tr)都有着明显的变化规律:对照CK(未修枝) < 中度修枝P1(修枝1/3) < 重度修枝P2(修枝2/3),即随着修枝程度的增加,蒸腾速率变大。对试验结果进行方差分析:修枝P跟未修枝CK之间有着显著性差异(P < 0.05),这表明修枝能够一定程度上提高树木叶片的蒸腾能力。树木自身存在着一种补偿调剂机制,修枝能过提高树木叶片蒸腾能力可能正是由于树木自身的这种补偿机制。林木为了补偿下部枝叶缺失而导致的总体水分吸收降低的不利影响,通过提高单叶蒸腾速率,提高林木吸水能力来进行补偿。

|
图 3 试验地Ⅰ(A)和试验地Ⅱ(B)生长季内生试验样木的Tr对比变化 Figure 3 The comparison of Tr of poplar at Site Ⅰ and Site Ⅱ during the growing season |
水势是反映树木水分状况的一个重要指标,也是代表树木抗旱生理特性的指标之一。叶水势反映叶片中水分子的能量水平,是水分运输的动力(王贵霞等,2003; 李华桢等,2006)。清晨叶水势(LWP)可以反映树木水分的恢复状况以及叶片的吸水能力,从而可以用来判断树木水分亏缺的程度及土壤水分供应的水平。这是由于树木进入夜间后,叶水势基本处于平缓稳定的恢复阶段,直到第2天清晨恢复到最高值。从图 4可知:试验样木叶水势都呈现明显的变化规律:对照CK(未修枝) < 中度修枝P1(修枝1/3) < 重度修枝P2(修枝2/3)。对试验结果进行方差分析:修枝P跟未修枝CK之间有着显著性差异(P < 0.05)。随着修枝强度的增强,叶水势绝对值减小,即叶水势变大,这说明树木修枝程度越强,树木越不亏缺水分,在相同土壤水分供应和生态环境的条件下,修枝使得树木的叶片恢复水分能力以及吸水能力得到提高。这种情况的出现,同样可能是基于树木自身补偿机制的调节作用,使树木最大可能的应对各种内在外在的变化。

|
图 4 生长季内4年(A)生和6年(B)生样木叶水势对比变化 Figure 4 The comparison of leaf water potential between 4-year-old and 6-year-old poplar during growing season |
1) 中度修枝P1 (修枝1/3)为较为合适的修枝强度。适当修枝P1,与正常月份未修枝对照CK相比,使树干液流量始终保持在较低的水平上,蒸腾速率以及叶水势等生理能力维持在合适的中间水平。P1修枝处理在一定程度上减小了杨树对水分的依赖程度,降低杨树的耗水量。陈森锟等(2008)的研究结果表明:P1修枝强度不仅不会损害到杨树的生长,而且还有着显著的增加材积生长量,减小木材的尖削度,提高木材利用率等作用。同时,较低水平的树干液流代表着树木对水需求量的降低,抗旱能力的提高,使林木更加容易度过极端天气状况,利于缓解林业用水量过大的问题。
重度修枝P2虽然使107杨的单叶蒸腾速率和叶水势等水分生理能力增强,但是其液流速率和液流通量大大降低,生长季内一直处于最低值状态。重度修枝P2损害林木的正常生长,明显减小林木的材积生长量(陈森锟,2008)。这可能是由于修掉大量的枝叶,林木的总体生理能力减弱,超过自身补偿的阈值,导致水分及营养等供应不足,损害林木的生长。
2) 修枝能使树木的蒸腾速率(Tr)、叶水势(LWP)生理能力得到明显的提高。目前认为这可能是基于树木自身的一种补偿调节机制。这种机制在国内外都有着相关研究,即林木为补偿下部枝叶缺失产生的不利影响,在一定条件下,通过提高单叶能力等途径进行补偿(Pinkard et al., 1999; Bayala et al., 2002; Schmidt et al., 2002; Neilsen et al., 2003; Bemiger et al., 2003; Burgess et al., 2004; Nabb et al., 2005; 王世绩等,1993)。
3) 从水分生理角度研究修枝对欧美杨107杨的影响,是为全面评估修枝对树木生长影响的重要内容。结合笔者实验室从材积生长量和相对尖削度等生长指标角度对修枝进行的试验研究,更为系统的为杨树修枝抚育技术提供理论基础和实践依据。修枝后树木水分生理指标的变化,材积生长量和相对尖削度等生长指标的变化,这可能是与树木自身的一种补偿机制有关,也可能与激素调节,源—库大小比等机制有关。这就需要研究人员进一步从宏观微观多角度来对修枝技术进行系统全面的研究,以得到更为全面的数据,使修枝作为基础性抚育措施得以真正的推广和应用。
修枝作为一种基础性林业抚育手段,在应用实践上的作用也需要进一步的深入研究。修枝过后,树木成熟后的材性和力学性质有何变化,这方面的研究对建筑业、木板家居业等行业有着重要影响; 修枝后树木耗水量减少,但是具体的灌水时间以及灌水量的确定仍需要林业工作者进一步的进行大量的试验研究测定。
陈森锟, 尹伟伦, 刘晓东. 2008. 修枝对欧美107杨木材生长量的短期影响[J]. 林业科学, 44(7): 130-135. DOI:10.11707/j.1001-7488.20080722 |
李华祯, 姚保强, 杨传强, 等. 2006. 4种经济树木水分生理及抗旱特性研究[J]. 山东林业科技, 163(2): 9-11. |
马履一, 王华田. 2002. 油松边材液流时空变化及其影响因子研究[J]. 北京林业大学学报, 24(3): 23-27. |
孙尚伟, 夏新莉, 刘晓东, 等. 2008. 修枝对复合农林系统内作物光合特性及生长的影响[J]. 生态学报, 28(7): 3185-3191. |
王保平, 李吉跃, 乔杰, 等. 2007. 修枝促接干对泡桐光合特性影响的研究[J]. 林业科学研究, 20(1): 119-124. |
王贵霞, 王海明, 李传荣. 2003. 五角枫金银木刺槐水分生理特性及抗旱性研究[J]. 山东林业科技, 5(3): 11-13. |
王华田, 马履一. 2002a. 油松、侧柏深秋边材木质部液流变化规律的研究[J]. 林业科学, 38(5): 31-37. |
王华田, 马履一. 2002b. 利用热扩式边材液流探针(TDP)测定树木整株蒸腾耗水量的研究[J]. 植物生态学报, 26(6): 661-667. |
王世绩, 刘雅荣. 1993. 杨树失叶对生长超越补偿作用的研究[J]. 林业科学研究, 36(3): 294-29. |
王华田, 赵文飞, 马履一. 2006. 侧柏树干边材液流的空间变化规律及其相关因子[J]. 林业科学, 42(7): 21-27. |
曾凡江, 张希明. 2002. 柽柳的水分生理特性研究进展[J]. 应用生态学报, 13(5): 611-614. |
张津林, 张志强, 查同刚, 等. 2006. 沙地杨树人工林生理生态特性[J]. 生态学报, 26(5): 1523-1532. |
Bayala J, Teklehaimanot Z, Quedraogo S J. 2002. Millet production under pruned tree crowns in a parkland system in Burkina Faso[J]. Agroforestry Systems, 54(3): 203-214. DOI:10.1023/A:1016058906682 |
Brodribb T J, Holbrook N, Gutiérrez M V. 2002. Hydraulic and photosynthetic co-ordination in seasonally dry tropical forest trees[J]. Plant Cell and Environ, 25(11): 1435-1444. DOI:10.1046/j.1365-3040.2002.00919.x |
Bemiger F, Salas E. 2003. Biomass dynamics of Erythrina lanceolata as influence by shoot-pruning intensity in Costa Rica[J]. Agroforestry Systems, 57: 19-28. DOI:10.1023/A:1022910310082 |
Gullo M. A., S. Salleo, M. A. LoGullo. 1992. Water storage in the wood and xylem cavitation in 1-year-old teigs of Populus deltoids Bartr[J]. Plant, Cell and Environment, 15(4): 431-438. DOI:10.1111/pce.1992.15.issue-4 |
Lawlor D W, Cornic G. 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants[J]. Plant Cell and Environ, 25(2): 275-294. DOI:10.1046/j.0016-8025.2001.00814.x |
Nabb K M, Vandersehaaf C. 2005. Growth of graded Sweetgum 3 years after root and shoot pruning[J]. New Forests, 29(3): 313-320. DOI:10.1007/s11056-004-5654-7 |
Neilsen W A, Pinkard E A. 2003. Effects of green pruning on growth of Pinus radiate[J]. Canadian Journal of Forest Research, 33(11): 2067-2073. DOI:10.1139/x03-131 |
Santiago L S, Goldstein G, Meinzer F C, et al. 2004. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees[J]. Oecologia, 140(4): 543-550. DOI:10.1007/s00442-004-1624-1 |
Schmidt T L, Wardle T D. 2002. Impact of pruning Eastern Redeedar (Juniperus virginiana)[J]. Wemon Journal of Applied Forestry, 17(4): 189-193. |
Pinkard E A, Battaglia M, Beadle C L, et al. 1999. Modeling the effect of physiological responses to green pruning on net biomass production[J]. Tree Physiology, 19(1): 1-12. DOI:10.1093/treephys/19.1.1 |