 2011, Vol. 47
2011, Vol. 47文章信息
- 杨东华, 赵雨森
- Yang Donghua, Zhao Yusen
- 不同生境对踏郎生长及土壤养分空间差异的影响
- Effects of Site Condition on Growth and Spatial Variations of Hedysarum Laeve
- 林业科学, 2011, 47(2): 180-186.
- Scientia Silvae Sinicae, 2011, 47(2): 180-186.
-
文章历史
- 收稿日期:2009-09-05
- 修回日期:2010-01-18
-
作者相关文章



踏郎(Hedysarum fruticosum),又称羊柴、山花子、山竹子、油葫芦稍,豆科岩黄芪属。踏郎是一种优良的保土固沙灌木,具有很强的根蘖能力,常“一株成林”,是优良的固沙造林植物种,大部分分布在我国内蒙和西北地区的固定、半固定沙丘上,而在内蒙古大面积的半流动沙地上和流动沙地的附近也有分布,用其固定沙丘(柏玉珍,1989)。踏郎的水土保持及经济效益都很高,自然生长带位于90°—125°E,38°—48°N之间,踏郎分布区的年降雨量一般为150~400 mm,干燥度为1.48~4.0(赵金荣,1985)。野生的踏郎多与木寥、沙篙、柠条等混生,组成复合群丛,有时也在半固定沙丘上形成大而集中的单独群丛。
不同生境对人工林生长及土壤养分的空间分布有着重要影响,且不同坡位土壤养分空间分布的差异主要是由坡面土壤性质和坡面养分在降雨侵蚀过程中的再分配所造成的(郭秀珍等,2005;钱凤军,2001)。植被对土壤养分也有重要影响,植物群落根系活动深度不同,对土壤养分的吸收强度和深度也不同(Stone et al., 1989;孟祥楠等,2007)。种群数量结构是种群的基本特征,种群的数量、年龄、密度、高度等特征是种群结构要素,是从不同角度对种群状况的反映。结合种群的生态学和生物学特征,研究人工林种群高度结构、径级结构不仅可以反映种群的现状,预测其未来,还可以展现植物种群与环境抗争的关系(Manuel et al.,2002;张文辉,1998)。本研究通过对人工林种群的径级和高度结构分析,来探讨人工防风固沙林踏郎种群不同生境条件下的生长情况(刘鑫等,2007)。本文选取6年生踏郎人工灌木防风固沙林为对象,研究了不同生境对踏郎人工林生长及土壤养分状况的影响,旨在揭示本地区地形条件与植物生长的关系,为该区水土保持措施的实施及生态环境建设提供理论依据。
1 研究区概况研究区位于赤峰市巴林右旗西北部的半流动沙地,地处大兴安岭东南缘,北与锡盟西乌珠穆沁旗交界,东与巴林左旗毗邻,西与林西县接壤,南与翁牛特旗隔西拉沐沦河相望,地理坐标为43°12′55″—44°27′52″N,118°12′09″—120°04′42″E,总面积9 638 km2,地处著名的科尔沁沙地腹地(孟琳等,2006)。据大板气象站统计资料,年平均气温4.9 ℃,1月平均气温-13.7 ℃,极端最低气温-31.1 ℃。7月平均气温22.2 ℃,极端最高气温38.7 ℃。≥0 ℃积温3 207.3 ℃,≥5 ℃积温3 061.6 ℃,≥10 ℃积温2 630.0 ℃。无霜期131天。年平均降水量350 mm,春季降水占全年总降水量的10%,夏季约占75%,秋季占12%,冬季只占全年的3%。年蒸发量为2 103 mm,全年蒸发量超过年降水量的6倍,干旱年份可达10倍以上。全年多为西、西北风,风势较大,年平均风速为3.4 m·s-1,全年大风日数平均为60天以上,最高年份达106天,多集中在4, 5月份。光照条件较好,太阳辐射总量为5 200~5 400 MJ·m2,生长季节(4—9月)的总辐射量占全年总辐射的65%。土壤以风沙土、棕壤土、栗钙土、草甸土和盐碱土为主,同时也有少量黑土(左小安等,2006;曹显军,1999)。在半流动沙地上的植物种类少,先锋植物为沙米(Agriophyllum squarrosum)、地锦草(Euphorbia humifusa )、北五加皮(Periplola sepium)等;继先锋草本之后,人工植物多为沙蒿(Artemisia arenaria)、柠条(Csragana intermedia)、踏郎和小叶杨(Populus simonii )等,沙地灌木、半灌木植被在沙丘上通过人工建植或天然落种方式存活下来。
2 研究方法 2.1 样地设置与外业调查在巴彦尔灯林场半流动沙丘梁地上选择不同生境(上坡、中坡、下坡)条件下6年踏郎作为研究对象,同时选取立地质量相近自然沙地(同坡面相邻平行50 m处)作为对照,分别设置20 m×20 m的样地各3块(自然沙地分别设置平行于上坡、中坡、下坡各3块样地,自然沙地土壤各层养分含量即为所设样地各层平均值,同时自然沙地地被层生长物种有:北五加皮、地锦草、冰草(Agropyron cristatum)、草地早熟禾(Poa pratensis)4个物种(Begon et al. 1990),于2008年6,7月进行野外调查。对样地内的踏郎进行了种群结构统计(踏郎高度每20 cm为一个高度级;踏郎人工林地径每1.5 cm为一个径级),按样地统计每一高度(径)级的个体数量,统计出踏郎人工林的高度(地径)结构;用米尺测量其高度、冠幅,以及采用游标卡尺测量其各株径级,同时统计株数(孟宪宇,1995;张文辉等,2002)。
2.2 土壤化学性质测定每个样地内分别在土壤剖面0~20 cm、20~40 cm、40~60 cm深处采集土样,室内风干,采用国家林业行业标准《森林土壤分析方法》测定土壤养分含量,包括土壤有机质、有效P、速效K、水解N、全N、全P和全K(关继义等,2005)。
3 结果与分析 3.1 不同生境踏郎人工林生长分析不同生境踏郎人工林平均树高、平均冠幅、平均株数、郁闭度、保存率及盖度均表现为下坡>上坡>中坡。这种情况在一定程度上是由于土壤养分在重力作用下体现出重力流失带而致,同时也由于生境条件的差异,下坡好于上坡,踏郎下坡生长最好,种群平均树高79.01 cm、平均株数64 567株·hm-2、郁闭度0.253、盖度为0.31、保存率达到82.43%;中坡踏郎生长最差, 种群平均树高68.12 cm、平均株数38 930株·hm-2、郁闭度0.10、盖度为0.13、保存率达到72.64%。出现这种现象的原因为:半流动沙丘在主风力作用下,下坡坡度较缓且植株密集受表面风力影响较弱,而中坡由于空气动力学及风沙物理学原理(王礼先等,2005),不仅受主风力的影响同时还受到上坡部分气体涡流作用,使得中坡沙地表面风速较大,进而导致沙地表面更干燥,导致植被生长条件很差,而上坡不断有沙粒沉积,使得表面的水分得以保护,地表层水分蒸发速率减少,生境条件好于中坡。表 2说明:半流动沙丘人工林踏郎平均地径为:上坡>下坡>中坡,说明沙埋对植株的地径影响较大,上坡和下坡沙埋效果较强。
|
|
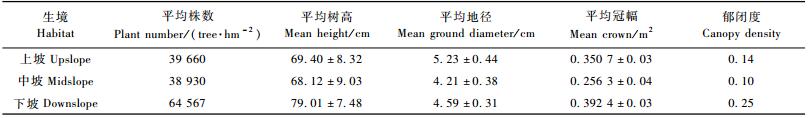
1) 不同生境踏郎种群径级结构 不同生境条件下踏郎固沙林的径级结构如图 1所示。在该地区不同生境条件下踏郎种群的平均地径出现差异,种群平均个体地径:上坡>下坡>中坡。上坡的踏郎种群径级主要分布在Ⅱ~Ⅵ径级,其中Ⅲ, Ⅳ径级最多,达到2 750株·hm-2,平均地径为5.23 cm;下坡的踏郎种群径级主要分布在Ⅱ~Ⅵ径级,其中Ⅲ径级最多,达到5 600株·hm-2,平均地径为4.59 cm;中坡的踏郎种群径级主要分布在Ⅱ~Ⅴ径级,其中Ⅲ径级最多,达到4 600株·hm-2,平均地径为4.21 cm。统计表明:踏郎径级主要分布在Ⅱ,Ⅲ,Ⅳ级,且占总径级79.4%。由此可知,不同的生境条件对人工林踏郎地径的影响很大。

|
图 1 不同生境条件下踏郎种群径级 Figure 1 Diameter class of Hedysarum Laeve population |
2) 不同生境踏郎种群高度结构 由图 2可以看出:在该地区不同生境条件下踏郎种群的平均高度级出现差异,种群平均个体高度:下坡>上坡>中坡。下坡的踏郎种群高度级主要分布在Ⅱ~Ⅷ级,其中Ⅲ高度级最多,达到4 100株·hm-2,平均树高为79.01 cm;上坡的踏郎种群高度级分布在Ⅰ~Ⅶ级,其中Ⅳ级最多,达到2 300株·hm-2,平均树高为69.40 cm;中坡的踏郎种群高度级主要分布在Ⅱ~Ⅶ级,其中Ⅲ级最多,达到3 600株·hm-2,平均树高为68.12 cm。统计表明:踏郎高度级主要分布在Ⅱ,Ⅲ,Ⅳ,Ⅴ级,占总高度级71.6%。由此可知,由于所处的生境条件不同,导致土壤养分、水分、光照等条件产生差异,下坡踏郎的平均树高最大。

|
图 2 不同生境条件下踏郎种群高度 Figure 2 Height of Hedysarum Laeve population |
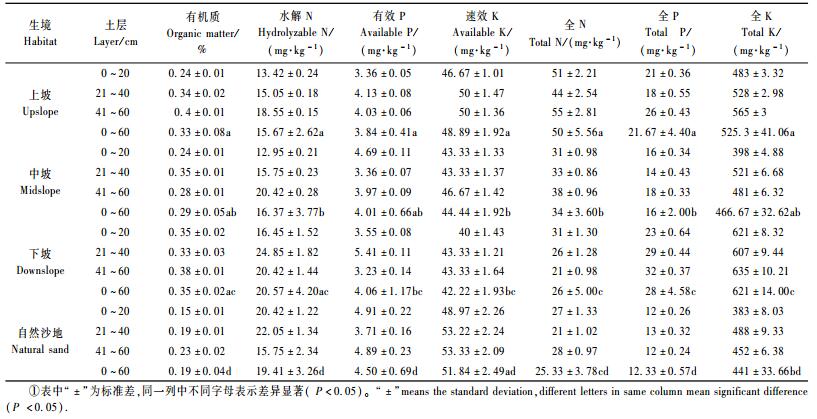
1) 不同生境踏郎人工林土壤化学性质组成特征 从表 3可知:同生境不同层间各自化学成分的标准差比较接近,在一定程度上验证了取样的准确性及样间误差在允许范围内,而不同生境间各化学成分用0~60 cm土层的平均值进行差异分析得出,不同生境相同化学成分间整体差异显著(P﹤0.05)。例如,土壤全P含量0~60 cm土层平均值差异显著(P﹤0.05),即上坡、中坡、下坡及自然沙地间差异分别为a, b, c和d代表。同时也可以从表中得出,上坡、中坡和下坡与自然沙地间差异更显著,这在一定程度上也体现出栽种人工林踏郎后极大地改良了土壤立地质量,有利于生态环境的恢复。
|
|
2) 不同生境踏郎人工林土壤有机质空间差异 踏郎人工林不同生境及自然沙地土壤有机质含量在0~60 cm土层中(除中坡)随土层增深而增加,且在50 cm土层均最大,按照上坡、下坡和自然沙地顺序则最大值分别为0.39%, 0.38%和0.23%,而中坡土壤有机质最大值则出现在30 cm土层,其最大值为0.35%,主要因为中坡植被郁闭度较小且土壤含水量较低,导致植被大部分根系分布在此深度,使得该层土壤有机质含量较高。另外,在0~20 cm土层中有机质(除下坡)含量均最小,按照上坡、中坡和自然沙地顺序最小值分别为0.23%,0.23%和0.16%,而下坡土壤有机质最小值则出现在30 cm土层,其最小值为0.33%,由于土壤养分在重力作用下体现出重力流失带且表层植被枯枝落叶归还量少、地表温度较高(土壤含水量较低)所致,而下坡部位各土层中平均有机质含量均极高,这与下坡的植被生长较好有关,而上坡和中坡则表现为交替变化,自然沙地各土层中有机质含量均最低,是由于地被植物少,枯落物归还量少,植被郁闭度很小,使得地表温度更高(土壤含水量极低)而致。土壤0~60 cm土层中有机质含量平均值为:下坡>上坡>中坡>自然沙地。

|
图 3 不同生境踏郎土壤有机质含量变化趋势 Figure 3 Organic matter of Hedysarum Laeve population |
3) 不同生境踏郎人工林土壤水解N空间差异 踏郎人工林不同生境及自然沙地土壤水解N含量在0~60 cm土层中的垂直变化分2种趋势:第一种趋势,上坡和中坡土壤水解N含量随土层增深而逐渐增加,且土壤水解N含量平均值较少,最大值均出现在50 cm土层中,分别为18.5和20.5 mg·kg-1,主要因为上坡和中坡土壤含水量较低,导致踏郎根系大部分生长于这一深度,同时制约土壤中无机态氮和部分有机质中易分解的、比较简单的有机态氮的释放,导致土壤水解N平均含量较低;第二种趋势,下坡和自然沙地土壤水解N含量随土层增深先增加后减少,且土壤水解N含量平均值较大,最大值均出现在30 cm土层,分别为25.2和22.3 mg·kg-1,由于下坡和自然沙地植被长势趋于稳定状态,土壤中含水量也较高,大多数植被根系在该层中能较好地加速土壤中无机态氮和部分有机质中易分解的、比较简单的有机态氮的释放,使得下坡和自然沙地土壤水解N平均含量较高且出现于该层土壤中,踏郎人工林不同生境土层中平均水解N含量总体表现为:下坡>自然沙地>中坡>上坡。

|
图 4 不同生境踏郎土壤水解N含量变化趋势 Figure 4 Hydrolytic N or Hedysarum Laeve population |
4) 不同生境踏郎人工林土壤有效P空间差异 踏郎人工林不同生境及自然沙地土壤有效P含量在0~60 cm土层中的垂直变化分2种趋势,上坡和下坡随土层增深先增加后减小趋势,主要因为30 cm土层分布一些不能充分利用微溶性、难溶性的磷酸盐化合物的植被根系,使得有效P含量偏高;而中坡和自然沙地则随土层增深先减小后增加趋势, 这是因为中坡和自然沙地植被郁闭度较小,使得地表风速较大,表面沙粒不断被吹走,进而土壤含水量较低,使得30 cm较深土层中分布植被根系较多,该土层中被植被根系吸收利用的土壤胶体表面吸附态和易交换态的磷含量较多,导致剩余有效P较少。土壤有效P含量不同土层中差异较大,但30 cm土壤深度为转折层,即上坡、下坡最大值出现于该层,分别为4.1,5.4 mg·kg-1;而中坡和自然沙地最小值出现于该层,分别为3.6和3.7 mg·kg-1。由图可知踏郎人工林不同生境土层中平均有效P含量总体表现为:下坡>自然沙地>中坡>上坡。

|
图 5 不同生境踏郎土壤有效P含量变化趋势 Figure 5 Available P of Hedysarum Laeve population |
5) 不同生境踏郎人工林土壤速效K空间差异 图 6表明:土壤中速效K含量在0~60 cm土层中(除中坡)随土层增深而先增加后减少趋势,且30 cm土层为土壤速效K富集层,按照上坡、下坡和自然沙地顺序则最大值分别为50.5,43.8和53.9 mg·kg-1;主要由于该层分布踏郎根系较少,直接吸收利用土壤中的交换性钾和水解性钾少,使得土壤中速效钾剩余较多;而中坡随土层增深而先减少后增加趋势,但在30 cm土层中出现最小值为43.1 mg·kg-1,是由于中坡30 cm土层分布踏郎根系较多,被根系吸收利用的速效K较多,所以最小值出现在该深度内。下坡平均速效K含量最低,这是因为下坡植被生长较好,对土壤中速效养分含量有较大的影响,已吸收利用土壤中的交换性钾和水解性钾较多,则剩余速效K含量较低;同理可知自然沙地平均速效K含量最大之因。踏郎不同生境速效K含量的最小值都出现在10 cm土层,这是由于表面10 cm土层中分布较多1至2年生的植被根系,而这些根系多数能较强吸收利用交换性钾和水解性钾。踏郎人工林不同生境土层中平均速效K含量总体表现为:自然沙地>上坡>中坡>下坡,同时也说明了土壤速效K含量与坡向呈正相关。

|
图 6 不同生境踏郎土壤速效K含量变化趋势 Figure 6 Available K of Hedysarum Laeve population |
6) 不同生境踏郎人工林土壤全N空间差异 图 7踏郎人工林不同生境及自然沙地土壤全N含量在0~60 cm土层中的垂直变化分2种趋势:一种趋势,下坡土壤全N含量随土层增深而减少,主要因为踏郎下坡根系大部分分布在50 cm土层,使得全N含量明显下降,最大值出现在10 cm土层,31 mg·kg-1; 另一种趋势,上坡、中坡和自然沙地土壤全N含量随土层增深而先减小后增加,且土壤全N含量平均值均较大,最大值均出现在50 cm土层中,分别为55,38和28 mg·kg-1,同时也可以得出土壤全N含量平均值总体表现为:上坡>中坡>下坡>自然沙地,主要因为由于不同生境条件土壤含水量影响,上坡根系分布明显最少,下坡根系较多,导致土壤全N含量较低。

|
图 7 不同生境踏郎土壤全N含量变化趋势 Figure 7 Soil total N of Hedysarum Laeve population |
7) 不同生境踏郎人工林土壤全P空间差异 图 8踏郎人工林不同生境土层中平均全P含量总体表现为:下坡>上坡>中坡>自然沙地。踏郎土壤全P含量在0~60 cm土层中的垂直变化下坡随土层增深逐渐增大;上坡和中坡先减少后增加;而自然沙地则体现出先增加后减少趋势,总体可以看出,除自然沙地最大值出现在30 cm土层,最大值为13 mg·kg-1;其余均出现在50 cm土层,按照上坡、中坡和下坡顺分别为26,18和32 mg·kg-1。自然沙地随土层增深先增加后减小趋势,主要因为30 cm土层分布一些不能充分利用微溶性、难溶性的磷酸盐化合物的植被(无踏郎)根系,使得全P含量偏高;而其他生境条件下踏郎根系在土壤50 cm土层有一定的分布,能保持一定的土壤含水量,有利于土壤中无机磷及交换态磷的释放,根系吸收利用的土壤胶体表面吸附态和易交换态的磷含量较多。

|
图 8 不同生境踏郎土壤全P含量变化趋势 Figure 8 Soil total P of Hedysarum Laeve population |
8) 不同生境踏郎人工林土壤全K空间差异 踏郎人工林土壤全K含量如图 9可知:中坡和自然沙地随土层增深而先增加后减少趋势,且30 cm土层为土壤全K富集层,最大值分别为521,488 mg·kg-1;上坡是逐渐增大、下坡则先减少后增加趋势。上坡和下坡土壤全K含量最大值出现在50 cm土层,主要由于上坡和下坡的踏郎长势较好,该层分布踏郎根系较多,直接吸收利用土壤中的交换性钾和水解性钾较多,同理可知中坡和自然沙地土壤中全K含量较少,踏郎人工林不同生境土层中平均全K含量总体表现为:下坡>上坡>中坡>自然沙地。

|
图 9 不同生境踏郎土壤全K含量变化趋势 Figure 9 Soil total K of Hedysarum Laeve population |
1) 不同生境对踏郎种群地径和高度的生长有显著影响,踏郎地径表现为:上坡>下坡>中坡;而踏郎高度则表现为下坡>上坡>中坡。另外,踏郎种群平均树高、平均冠幅、平均株数、郁闭度、盖度和保存率均表现为:下坡>上坡>中坡,踏郎下坡生长最好,从不同生境对踏郎种群生长影响可知,下坡长势最好,上坡优于中坡。
2) 踏郎下坡土层中有机质含量均较高,下坡生境条件对踏郎生长影响较大,对增加土壤有机质含量有重要的影响,而上坡和中坡部位有机质含量交替变化。不同生境土壤有机质含量呈波动式变化,土层40~60 cm有机质的含量均较大,这也是踏郎根最适宜生长的深度,而有机质含量的最小值均出现在0~20 cm土层中。在0~60 cm土层中(除中坡)随土层增深而增加,且在50 cm土层均最大,按照上坡、下坡和自然沙地顺序则最大值分别为0.39%, 0.38%和0.23%,而中坡土壤有机质最大值则出现在30 cm土层,其最大值为0.35%。
3) 从土壤养分空间差异整体可知,在0~20 cm土层范围内土壤有效P、速效K、水解N均是自然沙地含量最大;0~60 cm的平均速效K和水解N含量明显高于栽植有踏郎土层中平均速效K和水解N含量。踏郎土壤有效P、速效K和水解N的最大值均出现在30 cm的深度,分别为5.4,43.8和25.2 mg·kg-1,主要因为该土层是踏郎根系及地上其他植被根系分布过渡层,使得该层养分明显富集。而从土壤全N、全P和全K含量总体可知,土壤不同生境条件下有很大差异,这种影响主要由于生境不同导致,同时也能从中了解不同生境土壤立地质量,下坡好于上坡,最差为中坡。
4) 本次在巴彦尔灯林场研究表明,半流动沙丘栽种人工林踏郎固定后,其土壤得到极大改良,踏郎在生长过程中不但没有消耗沙土中极为有限的土壤养分,而且极大的改善了土壤的立地质量。不同生境条件对踏郎生长及土壤养分空间分布影响显著,是通过研究踏郎种群结构和土壤化学性质组成特征得出。
柏玉珍. 1989. 优良的水土保持树种——踏郎[J]. 水土保持科技情报, 1(1): 8-22. |
曹显军, 刘玉山, 斯钦昭日格. 1999. 踏郎、黄柳植物再生沙障治理高大流动沙丘技术的探讨[J]. 内蒙古林业科技, 1(1): 2-4. |
郭秀珍, 孙巍, 马占军. 2005. 踏郎沙地造林技术[J]. 内蒙古林业调查设计, 28(1): 1-2. |
关继义, 陈立新. 2005. 土壤实验实习教程[M]. 哈尔滨: 东北林业大学出版社: 85-187.
|
刘鑫, 满秀玲, 陈立明, 等. 2007. 坡位对小叶杨人工林生长及土壤养分空间差异的影响[J]. 水土保持学报, 10(5): 1-6. |
孟祥楠, 赵雨森. 2007. 嫩江沙地防风固沙林稳定性研究[J]. 东北林业大学林学院硕士学位论文. |
孟琳, 赵雨森. 2006. 科尔沁沙地植被沙障对环境因子改善作用研究[J]. 东北林业大学林学院硕士学位论文. |
孟宪宇. 1995. 测树学[M]. 哈尔滨: 北京:中国林业出版社: 18-116.
|
钱凤军. 2001. 踏郎固沙造林技术[J]. 林业科技通讯, 1(9): 1-3. |
王礼先, 王斌瑞, 朱金兆, 等. 2005. 林业生态工程学[M]. 2版. 哈尔滨: 北京:中国林业出版社: 59-188.
|
赵金荣. 1985. 踏郎[J]. 中国水土保持, 1(10): 1-3. |
左小安, 赵学勇, 赵哈林, 等. 2006. 科尔沁沙地草地退化过程中的物种组成及功能多样变化特征[J]. 生物多样性, 1(01): 5-10. |
张文辉, 赵则海, 孙海芹, 等. 2002. 东灵山辽东栎林优势林木种群直径结构的研究[J]. 植物研究, 22(1): 84-90. |
张文辉. 1998. 裂叶沙参种群生态学研究[M]. 哈尔滨: 东北林业大学出版社: 17-76.
|
Begon M, Harper J L, TownSend C R. 1990. Ecology:Individuals, Populations and Communities[J]. Boston:Blackwell Scientific Publications: 14-96. |
Manuel C, Molles J. 2002. Ecology, Concept and Applications[J]. New York:McGraw-Hill Companies: 186-254. |
Stone J A, Buttery B R. 1989. Nine forages and the aggregation of a clay loam soil[J]. Canadian Journal of Soil Science, 69: 16-169. |