 2011, Vol. 47
2011, Vol. 47文章信息
- 张志坚, 高健, 蔡春菊, 范少辉
- Zhang Zhijian, Gao Jian, Cai Chunju, Fan Shaohui
- 铅胁迫下菲白竹的矿质营养吸收和分配
- Absorption and Distribution of Mineral Nutrients in Pleioblastus fortunei under Lead Stress
- 林业科学, 2011, 47(1): 153-157.
- Scientia Silvae Sinicae, 2011, 47(1): 153-157.
-
文章历史
- 收稿日期:2010-01-13
- 修回日期:2010-11-07
-
作者相关文章



随着矿产、化工、冶金等工业排放的“三废”剧增以及汽车尾气排放量的增加,人类生存环境中的土壤、水体以及空气受重金属污染现象日趋严重,有些重金属通过食物链在人体内蓄积,严重危害人体健康(Kambhampati et al., 2003)。铅是一种常见的重金属,全球每年消耗量约为400万t,但仅有1/4得到回收利用,其余大部分都以不同形式进入环境引起污染(李建正,2006)。铅属于过渡元素,有d电子,而d电子在催化磁性方面有特殊效能,会对植物产生胁迫作用(李剑敏等,2007)。当胁迫强度超过植物的适应能力时,会影响植物对养分的吸收、转运、分配和代谢,严重时导致植物死亡(孙瑞莲等,2005)。但铅对植物矿质营养的作用规律依然不清楚(刘爱荣等,2009;Kopittke et al., 2007)。
菲白竹(Pleioblastus fortunei)是禾本科(Gramineae)竹亚科(Bambusoideae)苦竹属(Pleioblastus)植物。常被用于地被绿化或盆栽观赏,具有很强的适应能力,且耐修剪,病虫害极少发生(张春霞等,2006)。研究显示:低浓度汞胁迫下该竹种细胞膜透性、MDA含量、净光合速率、SOD活性、POD活性和游离脯氨酸含量均无明显变化,该竹种对汞胁迫有较强的抗性(汤玉香,2007)。但菲白竹是否抗铅胁迫及铅胁迫下菲白竹的生理响应机制尚无研究。由于铅在土壤中易于与有机质、氧化物、黏土矿物复合或吸附, 且易形成硫酸盐、磷酸盐和碳酸盐等无机化合物, 其生物活性降低, 从而限制了植物提取土壤中铅的能力,不利于研究有效铅对植物的伤害程度(何振立等,1998)。为避免此问题,本研究采用EDTA·Na2与Pb(NO3)2等比例螯合,测定菲白竹在不同浓度铅胁迫下各器官中矿质营养元素的含量,以了解铅胁迫对菲白竹矿质元素吸收和分配的影响,为菲白竹在土壤铅污染治理中的应用提供理论基础。
1 材料与方法试验材料为营养繁殖的2年生菲白竹苗,2009年4月引进到中国林业科学研究院温室盆栽,2009年8月开始对该竹种进行营养液水培。水培容器为80 cm×55 cm×12 cm的长方形塑料盒,盒内盛15 L营养液,用泡沫板浮于营养液面作为植株的固定支持物。
材料用1/2荷兰氏营养液培养10天,经观察生长正常后加入等量螯合的Pb(NO3)2与EDTA·Na2进行处理,根据预试验中菲白竹对铅的较强耐性,配制Pb浓度分别为0,828.8,1 657.6,2 486.4和3 315.2 mg·kg-1的营养液。另设对照,即营养液中不添加Pb(NO3)2。每个浓度处理2盒,每盒8丛,每丛10~15株,高约30 cm,生长状况良好且一致。铅胁迫处理11天后,3 315.2 mg·kg-1处理的菲白竹叶片开始枯黄萎蔫,经调制叶绿素荧光成像系统(IMAGING-PAM)测定,实际光能转化效率下降到0.3左右,表明光系统Ⅱ受到严重破坏。此时收获根系、地下鞭部、地上茎部和叶片,用蒸馏水和超纯水多次冲洗。
本试验采用电感耦合等离子体质谱仪(ICP-MS, 7500ce)法测定元素含量。该方法能一次测定样品中多种金属元素的含量,可避免用不同方法测定元素含量所引起的误差。所有样品在65 ℃下干燥48 h至恒质量,研磨成粉末,将粉末过40目尼龙筛。精确称取0.100 0 g样品,采用HNO3-H2O2体系消解(王艳泽等,2006)。测定样品中Pb,Na,K,P,Mg,Ca,Fe,Mn,Zn,Cu,B和Mo的含量,每个样品取3次重复。
生物富集又称生物浓缩,是生物有机体从周围环境中蓄积某种元素或难分解化合物,使生物有机体内该物质的浓度超过环境中浓度的现象,常用富集系数来表示。生物富集系数也称吸收系数,是指植物中某元素质量分数与土壤中元素质量分数之比;转运系数是地上部某元素质量分数与地下部某元素质量分数之比,用来评价植物将重金属从地下部向地上部的运输和富集能力。差异显著性分析和相关性分析由SPSS 13.0相应模块完成。
2 结果与分析 2.1 铅胁迫下菲白竹各器官矿质营养变化从表 1可见,铅对根系矿质营养的影响表现为: 1) 随着Pb浓度增大,Na含量增加,在2 486.4 mg·kg-1处理时达到峰值,Na含量与Pb浓度呈极显著正相关(0.876);2) 随着胁迫浓度增大,K含量逐渐下降,K与Pb浓度呈极显著负相关(-0.941);3) P与Pb浓度呈显著正相关(0.542);4) Ca,Mn,Fe和Zn在各胁迫下的含量低于对照;5) Cu含量在2 486.4 mg·kg-1铅胁迫处理下达到最大,随后降低;6) B和Mo含量较低,本试验中含量变化不明显。
|
|
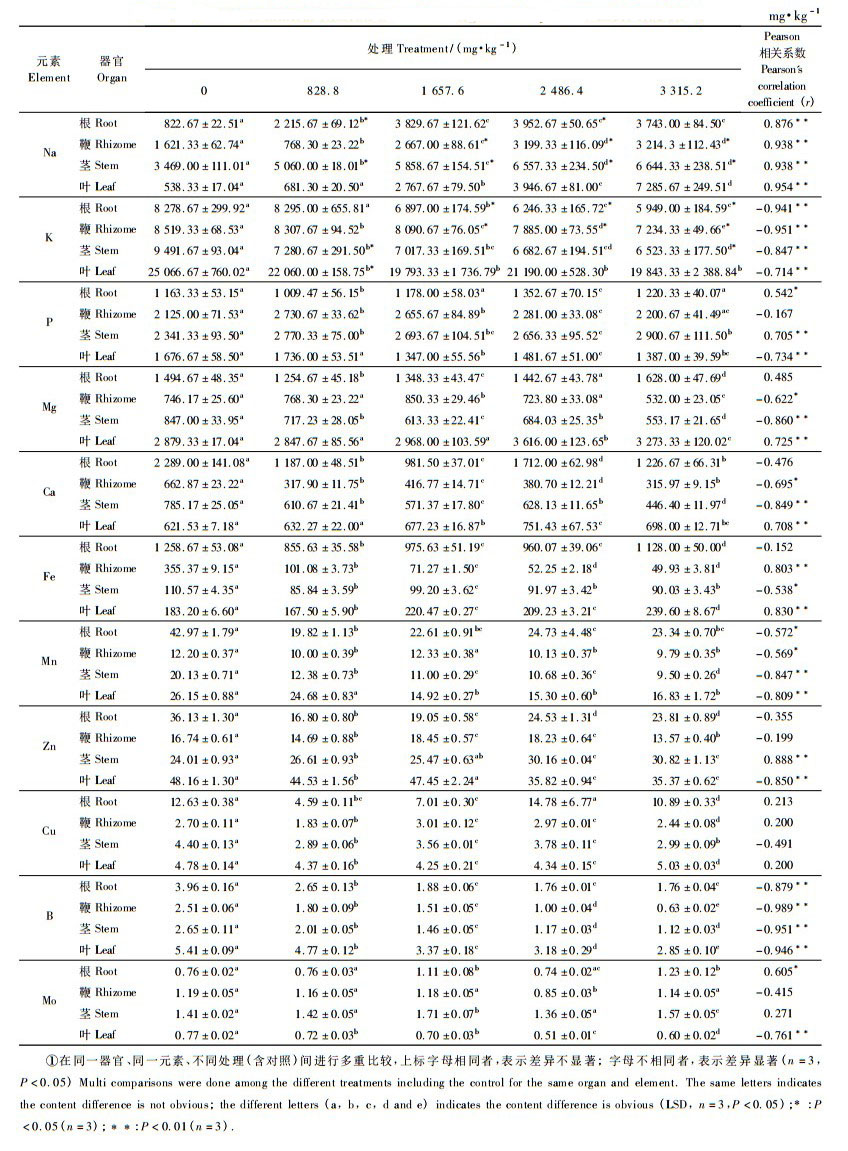
从表 1可见,铅对地下鞭部矿质营养的影响表现为: 1) Na含量随Pb浓度的增加而增加;2) K和Fe含量随Pb浓度的增加而下降;3) P和Mg在828.8和1 657.6 mg·kg-1处理下含量上升,在2 486.4和3 315.2 mg·kg-1处理下含量下降;4) Ca在各胁迫处理下的含量都低于对照;5) Mn和Zn在各处理下的含量差异不显著;6) B,Cu和Mo含量较低,变化也较小。
从表 1可见,铅对地上茎部矿质营养的影响表现为: 1) Na含量与Pb浓度呈极显著正相关(0.938);2) K,Mg,Ca和Mn含量与Pb浓度呈极显著负相关(相关系数分别为-0.847,-0.860,-0.849和-0.847);3) Fe含量与Pb浓度显著负相关(相关系数为-0.538);4) B,Cu和Mo含量较低,变化也较小。
从表 1可见,铅对叶片矿质营养的影响表现为: 1) Na,Mg,Ca和Fe含量与Pb浓度呈极显著正相关(相关系数分别为0.954,0.725,0.708和0.730);2) K,P,Mn和Zn含量随处理浓度的增加而下降;3) Cu,B和Mo含量较低,变化比较小。
所有铅胁迫处理下,在菲白竹的所有器官中Na含量均提高,K和B含量均下降,说明铅胁迫促进菲白竹对Na的吸收,同时抑制了菲白竹对K,B的吸收。对Mg而言,其在根系和叶片中含量上升,在鞭部和茎部含量有所下降。Ca在根系、鞭部和茎部呈下降趋势,但在叶片中呈上升趋势。在1 657.6和2 486.4 mg·kg-1处理下,Fe在叶片中的含量稳定增加,而在鞭部则下降。
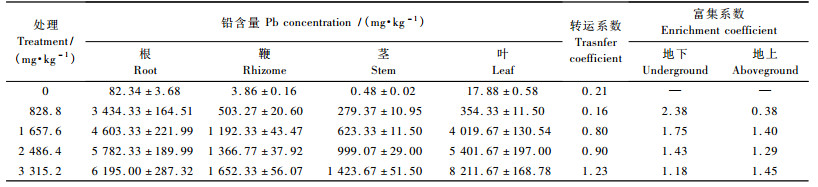
2.2 菲白竹各器官对铅元素的富集菲白竹各器官中铅含量也随着处理浓度的增加而逐渐升高,呈富集现象。处理后各器官铅含量如表 2所示。828. 8 mg·kg-1处理下Pb主要蓄积在菲白竹的地下部分,富集系数为2. 38,地上部分富集系数为0. 38,转运系数为0. 16。1 657. 6 mg·kg-1处理下菲白竹体内的铅元素蓄积量增加,转运系数迅速上升至0. 80,是828. 8 mg·kg-1浓度处理的5倍。在3 315. 2 mg·kg-1处理下Pb转运系数达到1. 23。
|
|
随着铅处理浓度的不断提高,菲白竹矿质元素含量也发生明显变化。菲白竹体内Pb和Na含量与培养液中Pb浓度呈显著正线性相关(根,鞭,茎和叶的相关系数分别为0. 876,0. 938,0. 938和0. 954),B和K含量随培养液中Pb浓度的增加而明显下降。所测元素中Na和K含量变化最明显; 同时,Mg,P,Ca和Fe等元素含量在不同器官也发生变化。正常的矿质营养平衡被打破,尤其是Na+/K+平衡被打破是铅毒害菲白竹的主要原因之一。植物抗逆性与Na+/K+离子平衡、Na+和K+的选择性吸收以及细胞原生质膜上Na+和K+的交换有重要关系(Niu et al., 1995)。Na+和K+选择性吸收的机制尚不清楚,可能与下列过程有关: 1) K+向内流动的选择性(流入选择性); 2) Na+和K+在(根皮层)原生质膜上的交换; 3) 液泡中Na+选择性积累及Na+和K+通过液泡膜的交换; 4) Na+和K+向导管中释放时的选择性; 5) 地上部枝叶对Na+的选择性吸收。近年来有一种假说,认为液泡在植物抗逆性方面起核心作用。Na+在液泡内的积聚与隔离和细胞质内K+的选择性吸收是渗透调节的关键因素(Sabirov et al., 1999)。胁迫条件下K+选择性地积累在细胞质中,而Na+则大量积聚在液泡中,在地上部分的枝叶部位,液泡中的Na+浓度有大幅度提高。所以,同样胁迫条件下,Na+能以更高的浓度在植株内积累。同时,植物体内Na+积累到一定程度时即对植物产生毒害,使植物体内自由基的含量增加,而细胞保护酶的活性受抑制,从而导致膜质的过氧化,增大了膜透性(柯玉琴等,1999),导致细胞外的盐离子大量进入细胞内,细胞内的溶质大量外渗,最终使得代谢紊乱,离子稳态被破坏,甚至导致植物死亡(王波等,2007)。K+是植物生长和发育必不可少的矿质离子,并在渗透调节、电中和、膜电位的调节和糖的协同运输等重要生理过程中起着至关重要的作用。在拟南芥(Arabidopsis thaliana)中发现K+吸收减少会引起叶片缺绿症(chlorosis)和生长抑制症状(Xu et al., 2006),这与本试验中钾离子缺失严重植株的表现症状相吻合。
本研究中K+含量的下降可能是K+的流失造成的,也可能与K+的吸收被阻碍有关,或二者兼有,其下降机理有待进一步探讨。铅胁迫下菲白竹叶片失绿发黄。当植物缺乏N,Mg,Fe,Mn,Cu,Zn和Mn等矿质元素中的一种时即不能形成叶绿素,易呈现缺绿症。本试验中菲白竹叶片中与叶绿素合成相关的Ca,Mg,Fe和Cu含量并未减少,而Mn和Zn含量下降,因此推测铅胁迫下的叶片失绿不是由于Ca,Mg,Fe和Cu的直接缺乏引起的,而可能与Zn和Mn的缺失有关。这点与镉胁迫下杨树叶片可能由Mn亏缺导致的黄化症和光合下降相符(万雪琴等,2009)。
菲白竹在Pb胁迫下可以有效富集Pb,根系和叶片是主要富集部位,富集量超过所谓“超富集”的参考值(1 000 mg·kg-1),在根系中最高可达到(6 195 ± 287. 32) mg·kg-1,在叶片中最高可达到(8 212 ± 168. 78) mg·kg-1。可见,菲白竹这种超富集植物可用于铅污染土壤的治理与修复。
根系是Pb进入植株体内的最初器官,根系对低浓度的Pb有着很强的吸收能力,将多数的Pb束缚在根里。在828. 8 mg·kg-1处理下,菲白竹根系Pb质量分数为(3 434 ± 164. 51) mg·kg-1,远超过鞭部的(503 ± 20. 60) mg·kg-1,茎部的(279 ± 10. 95) mg·kg-1和叶片的(354 ± 11. 50) mg·kg-1。随着铅浓度的增加,Pb在根系中的含量呈线性增长。在3 315. 2 mg·kg-1铅胁迫下,菲白竹根系Pb质量分数达到最高(6 195 ± 287. 32) mg·kg-1,远远超过超富集定义的参考值(1 000 mg·kg-1) (Liu et al., 2006)。Pb在菲白竹体内蓄积的规律是铅浓度≤ 2 486. 4 mg·kg-1时,根系>叶片>鞭部>茎部,铅浓度为3 315. 2 mg·kg-1时,叶片>根系>鞭部>茎部。
本试验中Mn含量在各器官中与Pb处理浓度呈显著负相关关系。在铅胁迫下Mn在菲白竹体内的含量变化,与NaCl胁迫下Mn在棉花(Gossypium hirsutum)幼苗叶片及根系中的含量变化(陈亚华等,2001)相类似,都是显著下降。在铅胁迫下Fe在菲白竹叶片中上升,与NaCl胁迫下Fe在棉花叶片中的含量变化趋势相似。但Zn在菲白竹叶片中下降,与NaCl胁迫下Zn在棉花叶片中的含量变化相反。可见,重金属胁迫与盐胁迫对植物体生长发育的影响在矿质元素水平上有相似之处,但由于处理材料和培养介质的不同,其离子含量变化也会有不同。各矿质元素之间存在一定的交互作用,交互作用的机理值得进一步探讨。本试验中铅处理浓度较高,有利于短时间内产生致死效应,但不利于研究低浓度铅胁迫下菲白竹的生理变化和矿质元素含量的关系,以后应开展低浓度处理试验。
陈亚华, 沈振国, 刘友良, 等. 2001. NaCl胁迫下棉花幼苗的离子平衡[J]. 棉花学报, 13(4): 225-229. |
何振立, 周启星, 谢正苗. 1998. 污染及有益元素的土壤化学平衡[M]. 北京: 中国环境科学出版社: 129-303.
|
刘爱荣, 张远兵, 张雪平, 等. 2009. 铅污染对高羊茅生长、无机离子分布和铅积累量的影响[J]. 核农学报, 23(1): 128-133. |
李剑敏, 杨劲松, 杨晓英, 等. 2007. EDTA对铅污染土壤上芥菜生长及铅积累特性的影响[J]. 土壤通报, 38(6): 1178-1181. |
李建正. 2006. 环境毒理学[M]. 北京: 化学工业出版社: 135.
|
柯玉琴, 潘延国. 1999. NaCl胁迫对甘薯叶片叶绿体超微结构及一些酶活性的影响[J]. 植物生理学报, 25(3): 229-233. |
孙瑞莲, 周启星. 2005. 高等植物重金属耐性与超积累特性及其分子机理研究[J]. 植物生态学报, 29(3): 497-502. DOI:10.17521/cjpe.2005.0066 |
汤玉香. 2007. 三种观赏竹对汞胁迫的生理响应及硅和稀土的调控作用. 南京林业大学硕士学位论文.
|
万雪琴, 张帆, 夏新莉, 等. 2009. 镉处理对杨树矿质营养吸收和分配的影响[J]. 林业科学, 45(7): 45-51. DOI:10.11707/j.1001-7488.20090708 |
王波, 宋凤斌, 张金才. 2007. 植物耐盐性研究进展[J]. 农业系统科学与综合研究, 23(2): 212-216. |
王艳泽, 施燕支, 王英锋, 等. 2006. ICP-MS对土壤和灌溉水样品中部分微量元素的测定[J]. 首都师范大学学报: 46-49. DOI:10.3969/j.issn.1004-9398.2006.02.012 |
张春霞, 王福升, 黄月英. 2006. 菲白竹组培繁殖技术研究[J]. 林业科技开发, 20(5): 31-33. |
Liu S C, Xiao L T, Liao Baihan, et al. 2006. Effects of lead stress on antioxidative enzyme activities and isoenzymes in cucumber seedlings[J]. Chinese Journal of Applied Ecology, 17(2): 300-304. |
Kambhampati M S, Begonia G B, Begonia M F T, et al. 2003. Phytoremediation of a lead-contaminated soil using morning glory (Ipomoea iacunosa):effects of a synthetic chelate[J]. Environmental Contamination and Toxicology, 71(2): 379-386. DOI:10.1007/s00128-003-0175-1 |
Kopittke P M, Asher C J, Blamey F P C, et al. 2007. Toxic effects of Pb2+ on the growth and mineral nutrition of signal grass(Brachiaria decumbens) and rhodes grass(Chloris gayana)[J]. Plant and Soil, 300(1/2): 127-136. |
Niu X, Bressan R A, Hasegawa P M, et al. 1995. Ion homeostasis in NaCl stress environments[J]. Plant Physiology, 109(3): 735-742. DOI:10.1104/pp.109.3.735 |
Sabirov R Z, Azimov R R, Ando-Akatsuka Y, et al. 1999. Na+ sensitivity of ROMK1 K+ channel: role of the Na+/H+ antiporter[J]. Journal of Membrane Biology, 172(1): 67-76. DOI:10.1007/s002329900584 |
Xu J, Li H D, Chen L Q, et al. 2006. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis[J]. Cell, 125(7): 1347-1360. DOI:10.1016/j.cell.2006.06.011 |