 2011, Vol. 47
2011, Vol. 47文章信息
- 陈在新, 潘娟, 江道菊
- Chen Zaixin, Pan Juan, Jiang Daoju
- 不同成熟期板栗品种(系)种子发育期主要营养物质与内源激素含量的动态变化
- Dynamic Changes of Main Nutrients and Endogenous Hormones in Seed of Different Mature Period Chinese Chestnuts During their Seed Development
- 林业科学, 2011, 47(1): 73-78.
- Scientia Silvae Sinicae, 2011, 47(1): 73-78.
-
文章历史
- 收稿日期:2010-01-22
- 修回日期:2010-03-27
-
作者相关文章


板栗(Castanea mollissima)属于壳斗科(Fagaceae)栗属(Castanea)坚果类植物。中国板栗栽培历史悠久,品种资源丰富。我国栽培的板栗品种成熟期较为集中、上市期拥挤,中晚熟品种坚果虫害严重,而且贮藏条件要求高、成本高,已成为板栗果实综合开发的最大障碍(席客等,1996; 鲁周民等,2003; 范伟国等,2007)。因此,生产上调控坚果成熟期,提早应市,延长供应期,满足市场需求; 缩短贮藏时间,减轻食果害虫危害,提高商品率和经济效益十分必要。目前,果树发育期内源激素含量的变化在多种果树上已有研究报道(李秀菊等,2000; 樊卫国等,2004; 刘旭,2008),板栗种子发育过程中营养物质与内源激素含量的变化研究却很少(周志翔,1999; 周志翔等,2002; 曾柏全等,2006),且研究的仅为单一品种,多个品种的系统研究尚为空白。本研究旨在通过比较多个不同成熟期的板栗品种种子整个发育期的主要营养物质与内源激素含量的动态变化,掌握其变化的特点和规律,探讨板栗种子成熟机制,为有效调控种子成熟期提供理论依据,并为采用分子生物学手段改良品种奠定生理学基础。
1 材料与方法 1.1 试验材料2008年取材于荆州松滋市陈店镇板栗资源圃,试材为以栽培品种实生苗为砧木嫁接的0212,0214,0204,02064个在板栗良种选育中处于中试中的品系和新品种鄂栗1号、鄂栗2号(以下简称鄂1、鄂2),6个品系(种)经济性状稳定、优良。0212、鄂2、0214称为早熟品种(系)(8月中旬成熟),鄂1、0206称为中熟品种(系)(八月下旬成熟),0204称为晚熟品系(九月中旬成熟)。各品种(系)选取生长结果良好的8年生树15株,5株小区,3次重复。以雌花柱头反卷45°前1天为花后第0天,记为0天。于花后第15天(幼胚开始发育)至完熟时(刺苞1/3出现开裂),每10天采样1次。每次每株随机取总苞(种子)6枚,每个品种(系)共30枚(前期多取),用冰盒带回。鲜样带回后立即测定乙烯释放量和Vc含量,用于其他内源激素及营养物质含量测定的样本液氮速冻2~3 min后,于-70 ℃低温保存待测。各样品同一指标重复测定3次。
1.2 试验方法内源激素IAA,GA3,ABA,ZT,Eth含量均采用气相色谱法(杜黎明等,2000; 刘志勇等,2006)测定,使用日本岛津GC-14C型气相色谱仪,FID检测器(幼果期Eth测定除去刺苞,未伤及幼嫩种子)。营养物质淀粉、可溶性总糖(以下简称总糖)含量采用蒽酮比色法(张友杰,1980)(干样)、Vc含量采用2,6-二氯酚靛酚法(鲜样)、可溶性蛋白质(以下简称蛋白质)含量采用考马斯亮兰G-250法(李合生,2000)(干样)测定。各样品同一指标重复测定3次。
1.3 数据分析试验数据使用DPS软件进行统计处理,SSR法多重比较,逐步回归法回归分析(唐启义等,2007)。
2 结果与分析 2.1 不同成熟期板栗品种(系)种子发育期主要营养物质含量的变化6品种(系)种子发育期主要营养物质含量的动态变化如图 1A~D所示。图 1A表明:淀粉含量变化均呈现“升-降-升”的规律,有2次高峰期。早、中熟品种(系)花后25天出现1个峰值,晚熟品系0204晚10天出现。完熟前10天左右各品种出现第2次峰值,峰值均极显著高于第1次(P < 0.05)。总体上看,在整个发育期淀粉含量早熟品种(系)>中熟品种(系)>晚熟品系,且均在完熟前20~30天开始大量积累。完熟时鄂2的淀粉含量最大为65.29%,极显著高于其他品种(系)(P < 0.01),0212, 0214显著高于0206, 0204,其他品种(系)间差异不显著。

|
图 1 不同成熟期板栗品种种子发育期营养物质动态变化 Figure 1 Changes of main nutrients during seeds development of different chestnuts |
总糖含量变化基本呈现“慢-快”的上升趋势(图 1B)。不同成熟期的品种(系)初期(花后20~30天)变化均较平稳,基本保持一定含量,完熟前20~30天开始大量积累。完熟时鄂2的总糖积累量最大为17.48%,显著高于晚熟品系0204(P < 0.05),其他品种(系)间差异不显著。整体上看,总糖含量在发育中、后期早熟品种(系)>中熟品种(系)>晚熟品系。
种子发育期蛋白质含量呈双“S”型变化(图 1C)。花后10天及完熟前10天左右含量均较高,且两峰值早熟品种(系)>中熟品种(系)>晚熟品系,但完熟时品种(系)间含量差异不显著。
种子发育期Vc含量呈线性上升趋势(图 1D)。含量与花后天数作线性回归分析,r分别为0.964 3(0212,F=53.06),0.967 4(鄂2, F=58.41),0.928 1(0214, F=24.86),0.939 4(鄂1, F=37.53),0.954 7(0206, F=51.46)、0.951 1(0204, F=66.30),表明各品种(系)Vc含量与花后天数分别呈极显著线性正相关。完熟时Vc含量最高,其中鄂2显著大于0204和0206(P < 0.05),其他品种(系)间差异不显著。
2.2 不同成熟期板栗品种(系)种子发育期内源激素含量的变化6品种(系)种子发育期内源激素含量的变化如图 2A~D所示。IAA含量呈“降-升-降”的变化趋势(图 2A)。花后15天各品种含量均最高,早、中熟品种(系)于花后45天,晚熟品系于花后55天开始递减,直至完熟时均降到最低。对各品种(系)种子完熟时IAA含量与其发育天数作线性回归分析F=4.689 67,结果线性相关不显著。完熟时IAA含量除0204,0206显著高于0212、鄂2、0214、鄂1(P < 0.05) 外,其他品种(系)间差异不显著。

|
图 2 不同成熟期板栗品种种子发育期激素含量动态变化 Figure 2 Changes of endogenous harmones during seeds development of different chestnuts |
6品种(系)ZT含量呈“单峰”变化(图 2B)。初期含量均较高,早熟品种(系)于花后25天,中、晚熟品种(系)于花后35天开始递减,至完熟时均降到最低。中、晚熟品种(系)递减期比早熟品种(系)推迟10天。对各品种(系)种子完熟时ZT含量与相应发育天数作线性回归分析,F=0.382 8,线性相关不显著; 多重比较结果,品种(系)间含量差异也不显著。
6个品种(系)种子内GA3含量变化差异较大(图 2C)。早、中熟品种(系)种子呈单峰曲线变化,于花后35天出现高峰; 晚熟品种0204种子呈双峰曲线变化,分别于花后25天和花后65天出现一大一小2个峰值,花后65天后逐渐降低至完熟时。对完熟时各品种(系)GA3含量与相应发育天数作线性回归分析,F=0.854 06,结果线性相关不显著,但0212,0206,鄂1,0204的含量显著低于鄂2,0214(P < 0.05),其他品种(系)间差异不显著。
ABA含量呈现波幅较小的“升-降-升”的变化过程(图 2D)。ABA含量早、中、晚熟品种(系)均于花后15天开始小幅度增加,早熟品种(系)于花后55天、中熟品种(系)于花后65天、晚熟品系于花后85天开始较大幅度的增加,直至种子完熟时达到最高值。开始较大幅度增加的时期,中、晚熟品种(系)比早熟品种(系)分别推迟10,30天。对成熟时ABA含量与种子发育天数作线性回归分析,F=14.438 47,r为-0.884 91,可知其呈显著线性负相关。总体上看,整个发育期ABA后期含量大于前期含量,其中,鄂2花后65天达到最高值0.161 μL·g-1。多重比较结果表明:早熟品种鄂2极显著高于、早熟品系0212,0214显著高于中晚熟品种(系)0204,0206和鄂1。
花后35天所有品种(系)种子中Eth开始产生,早、中、晚熟品种(系)分别于花后55,65,85天急剧增加,10天后(完熟时)达到最高值(图 3)。其中早熟的3个品种(系)Eth释放量极显著高于中、晚熟的0206和0204(P < 0.01),鄂2、0214极显著高于鄂1(P < 0.01)。对完熟时Eth释放量与种子发育天数作线性回归分析,F=22.105 19,r为-0.920 20,二者呈极显著线性负相关。

|
图 3 不同熟期板栗品种种子发育期Eth含量动态变化 Figure 3 Changes of Eth contents during seeds development of different chestnuts |
各品种(系)中种子发育期营养物质含量与内源激素含量间的通径分析结果如表 1所示。淀粉含量(Y1) 与ABA的含量呈极显著正相关,与IAA呈极显著负相关,与GA3呈显著负相关(表 1)。淀粉含量与内源激素含量有极显著的线性相关(F=26.020 3),决定系数为0.788 01,表明在Y1的总变化中,由变量X(1~5) 线性说明的部分占78.801%。X1的直接通径系数为-0.837 81,绝对值最大,因而IAA, ABA含量在淀粉含量变化中起主要作用,IAA的作用比其他激素更大,ABA次之。
|
|
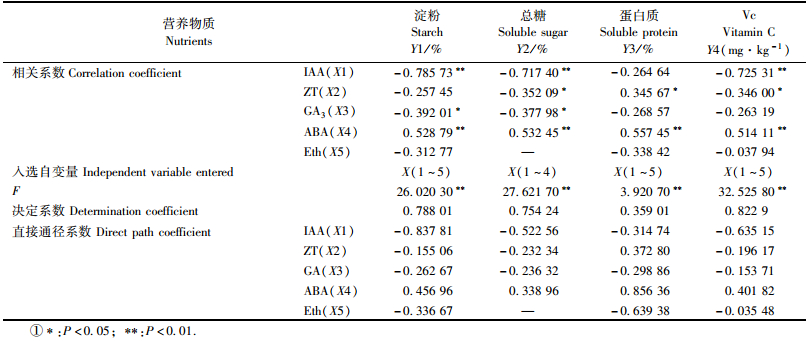
种子中总糖含量(Y2) 与ABA的含量呈极显著正相关,与ZT呈显著负相关。在Y2的总变化中,由变量X(1~4) 线性说明的部分占75.424%(F=27.621 7,决定系数为0.754 24),IAA的直接通径系数绝对值最大,ABA次之。总糖含量和淀粉含量与内源激素含量的线性关系基本相同。
蛋白质含量(Y3) 与ABA的含量呈极显著正相关,与ZT含量呈显著正相关。在Y3的总变化中,由变量X(1~5) 线性说明的部分占35.901%(F=3.920 7,决定系数为0.359 01)。ABA的直接通径系数绝对值最大,Eth次之。
Vc含量(Y4) 与ABA的含量呈极显著正相关,与IAA呈极显著负相关,与ZT呈显著负相关。在Y4的总变化中,由变量X(1~5) 线性说明的部分占82.29%(F=32.525 8,决定系数为0.822 9)。IAA的直接通径系数绝对值最大,ABA次之。
综上所述,各品种种子中IAA,ZT,GA3含量的减少与ABA含量的增加有利于种子中淀粉、总糖、蛋白质、Vc的积累,其中IAA,ABA对各营养物质的积累影响最大(以IAA的作用更大)。有研究认为,薄壳油栗胚胎发育期间(不同于本文研究的种子发育期)内源激素GA3含量的增加,ABA含量的降低,有利于子房还原糖、淀粉、蛋白质的积累(周志翔等,2002)。本研究与其结论不一致,可能是由于试材品种间遗传因子及研究的种子发育阶段不一致所致。
3 结论与讨论 3.1 板栗品种(系)种子发育期各种营养物质含量的变化趋势相似6个板栗品种(系)种子发育期淀粉含量呈现“升-降-升”的变化趋势,蛋白质含量呈双“S”型变化,总糖含量呈“慢-快”的上升趋势,Vc含量与种子发育天数均呈现极显著线性相关关系。
不同成熟期品种(系)种子在发育初期淀粉积累量少,转化成总糖的也少,随着种子的发育淀粉含量不断增加,成熟前20~30天开始大量积累,总糖含量也大量增加; Vc含量随着果实的发育逐渐加大积累量。
各品种(系)种子生育期分别为60天(鄂2)、62天(0214)、65天(0212)、73天(鄂1)、75天(0206)、94天(0204)。整个生育期或大部分时期,淀粉、总糖、蛋白质、Vc含量均呈现早熟品种(系)>中熟品种(系)>晚熟品系的现象,尤其是早熟品种鄂2,生育期短,营养物质积累也快。
3.2 板栗种子发育期内源激素含量的变化是引起种子中营养物质含量变化的重要原因板栗种子发育过程中,内源激素IAA,ZT,GA3呈递减趋势,ABA,Eth呈递增趋势。有研究表明:IAA,ZT,GA3能促进果实细胞分裂和体积膨大,ABA和Eth能启动成熟相关酶的合成,促进营养物质的相互转化与积累(于萍,1994)。本研究中板栗种子发育初期IAA,ZT,GA3含量高,故总苞逐渐膨大,种子中营养物质淀粉、总糖、Vc含量缓慢增长。随着IAA,ZT,GA3含量的减少,ABA和Eth的逐渐增加,种子由膨大变化转向营养物质的积累和转化,故淀粉、总糖在成熟前快速增加,同时淀粉部分转化成糖类。板栗种子发育过程中IAA,ZT,GA3含量的逐渐减少,ABA和Eth含量的逐渐增加有利于淀粉、总糖、蛋白质和Vc的积累。
3.3 种子发育期内源激素变化的起始与持续时间的差异是导致板栗品种成熟期不同的重要原因IAA含量早、中熟品种(系)于花后45天开始递减至成熟时,晚熟品种系推迟10天,分别持续20,30,40天; ZT含量早熟品种(系)于花后25天开始递减至成熟熟时,中、晚熟品种(系)均迟10天,分别持续40,40,60天; GA3含量早、中熟品种(系)于花后35天开始递减至成熟时,晚熟品系推迟30天,分别持续30,40,30天。ABA,Eth含量早、中、晚熟品种(系)分别于花后55,65,85天开始急增至成熟时,中、晚熟品种(系)比早熟品种(系)分别推迟10,30天,均持续10天。
坚果成熟时各品种(系)IAA,GA3,ZT含量分别与其发育天数作线性回归分析,线性相关均不显著; 成熟时IAA含量0204,0206显著高于0212,鄂2,0214,鄂1,其他品种(系)间差异不显著; ZT含量品种(系)间差异也不显著; GA3含量鄂2,0214显著高于0206,鄂1,0204,0212,其他品种间(系)差异不显著。坚果成熟时各品种(系)ABA,Eth含量分别与其发育天数的线性回归分析结果表明:ABA,Eth与坚果发育天数分别呈显著、极显著线性负相关。
综上分析可知:IAA,GA3,ZT,ABA,Eth在坚果发育期变化的起始时间及持续时间影响着板栗种子的成熟。各品种(系)种子中IAA,GA3,ZT含量在成熟前均开始减少,但起始时间不同,持续时间也不同,基本上晚熟品系持续时间长于中熟品种(系),中熟品种(系)长于早熟品种(系); 而各品种(系)种子中ABA,Eth含量在成熟前均迅速增加,但起始时间不同,持续时间相同,早熟品种(系)起始时间早于中熟品种(系),中熟品种(系)早于晚熟品系。故成熟前种子中IAA,GA3,ZT含量开始减少的时间与ABA,Eth含量开始增加的时间,以及各自持续时间的不同是引起6个板栗品种(系)间成熟期不同的重要原因。IAA,GA3,ZT含量开始减少的时间越早,种子膨大期持续时间越短,ABA,Eth含量开始增加的时间越早,成熟期就越早。反之,成熟期越晚。这与刘旭(2008)的研究结果一致。生产中可依据此原理和种子发育过程中IAA,ZT,GA3含量的逐渐减少,ABA和Eth含量的逐渐增加有利于淀粉、总糖、蛋白质和Vc的积累的规律,综合考虑对品种成熟期、产量和品质的要求,合理施用IAA,GA3,ZT和ABA,Eth 2类生长调节剂来调节果实成熟期,提高果实产量和品质。
由于植物体内激素种类较多,对果实发育的作用既不相同又相互协调(Yuan et al., 1988; Kojima et al., 1994),因此生长调节剂在板栗种子发育的不同阶段对内源激素的平衡调节作用及效应还有待于进一步研究。
杜黎明, 许庆琴. 2000. 气相色谱法分离和测定3种植物内源激素[J]. 色谱, 18(1): 67-69. |
樊卫国, 安华明, 刘国琴, 等. 2004. 刺梨果实与种子内源激素含量变化及其与果实发育的关系[J]. 中国农业科学, 37(5): 728-733. |
范伟国, 李玲, 刘树增. 2007. 板栗标准化安全生产[M]. 北京: 中国农业出版社: 221-222.
|
李合生. 2000. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社: 124.
|
李秀菊, 刘用生, 束怀瑞. 2000. 不同成熟型苹果果实生长发育过程中几种内源植物激素含量变化[J]. 植物生理学通讯, 36(1): 7-10. |
刘旭. 2008. 成熟期不同的梨品种果实生长发育机理探讨. 四川农业大学博士学位论文.
|
刘志勇, 沈春章, 董元彦. 2006. 气相色谱法速测油菜中的乙烯释放量[J]. 化学与生物工程, 23(2): 55-56. |
鲁周民, 李文华, 张忠良, 等. 2003. 采收成熟度对板栗品质及贮藏性的影响[J]. 食品科学, 24(7): 153-155. |
唐启义, 冯明光. 2007. DPS数据处理系统:实验设计、统计分析及数据挖掘[M]. 北京: 科学出版社: 636.
|
席客, 徐福元, 戴雨生. 1996. 板栗球果生命表的研究[J]. 江苏林业科技, 23(4): 6-10. |
于萍. 1994. 果实成熟机理和几个生理问题[J]. 生物学杂志, 4: 9-11. |
曾柏全, 陈建华, 姚跃飞. 2006. 板栗胚胎发育期间内源激素的动态变化[J]. 河南农业科学, 6: 95-97. DOI:10.3969/j.issn.1004-3268.2006.02.027 |
张友杰. 1980. 蒽酮分光光度法测定果蔬中的葡萄糖、果糖、蔗糖和淀粉[J]. 分析化学, 5(3): 167-171. |
周志翔. 1999. 板泉空苞形成与调节的生理机制研究. 华中农业大学博士学位论文.
|
周志翔, 王鹏程, 徐永荣, 等. 2002. 板栗子房内源激素与主要营养物质含量变化及其关系研究[J]. 林业科学, 38(3): 167-170. DOI:10.11707/j.1001-7488.20020329 |
Kojima K, Yamada Y, Yamamoto M. 1994. Distribution of ABA and IAA, within a developing valencia orange fruit and its parts[J]. J Japan Soc Hort Sci, 63(2): 335-339. DOI:10.2503/jjshs.63.335 |
Yuan R C, Huang H B. 1988. Litchi fruit abscission: its pattern, effects of shading and relation to abscsic acid[J]. Scientia Horticulturae, 36(3/4): 281-292. |