 2011, Vol. 47
2011, Vol. 47文章信息
- 邢世岩, 李际红, 邢浩, 张芳, 韩晨静, 唐海霞
- Xing Shiyan, Li Jihong, Xing Hao, Zhang Fang, Han Chenjing, Tang Haixia
- 叶籽银杏种实形态解剖特征比较
- Comparative Studies on Morphology of Seeds of Ginkgo biloba var. epiphylla
- 林业科学, 2011, 47(1): 56-61.
- Scientia Silvae Sinicae, 2011, 47(1): 56-61.
-
文章历史
- 收稿日期:2009-10-12
- 修回日期:2010-03-31
-
作者相关文章


2. 山东中医药大学 济南 250355
2. Shandong University of Traditional Chinese Medicine Jinan 250355
银杏(Ginkgo biloba)系最古老的种子植物之一,从系统发育的角度上看,银杏被认为是一种介于苏铁(Cycas revoluta)和其他种子植物间的独特的类群(Burleigh et al., 2004)。银杏具有明显的3层珠被(福斯特等,1983; Favre-Duchartre,1958); 配子体(gametophyte)发育经历含大量游离核的阶段; 没有胚珠苞片(ovule bract)和特殊的球果状大孢子叶球(Douglas et al., 2007; 邢世岩等,1996)。对不同银杏种质的正常种实形态、生理指标的研究已有报道(邢世岩等,2001; 2005;张云跃等,2001)。1891年,Shirai(1891)最早发现了叶籽银杏,但是直到1927年日本植物分类学家Makino首次把叶籽银杏定为变种:Ginkgo biloba L. var. epiphylla Mak.,并被后人引用(吉冈金市,1967)。先后有Fujii(1896)、Sakisaka(1927)、松田孫治(1932)、吉冈金市(1967)、Soma(1999)等学者对叶籽银杏的种实进行了描述,但均不系统。作者所在课题组对叶籽银杏的形态发育、叶解剖、核型及matK和ITS序列等进行了报道(李士美等,2007; 邢世岩等,2007; 李保进,2008; 李保进等,2007)。正常胚珠在树体上处于短枝顶端站立位,而叶籽银杏胚珠的着生方式为叶生,叶缘或叶面着生,每叶“胚珠”1~15个不等,常有1个成熟,稀2(3~4) 个成熟。一叶生一种实者,叶生种实着生处多偏离叶子正中或生于叶缘一侧; 而一叶具有2种实者,叶生种实多着生在叶片横长的1/3处左右,一般2个种实形状、大小、颜色均相似,稀一大一小; 一叶生3种实者,叶生种实多着生于叶片横长1/4处(Sakisaka,1927; 李士美等,2007)。
本研究采用作者等在山东境内发现的8株叶籽银杏(这是目前世界上唯一在同一个区域发现最多的叶籽银杏)上的正常种实和叶生种实为试材,对其种实形态多样性进行比较研究,并对其胚胎学意义进行了探讨,以期为叶籽银杏的系统发育提供依据。
1 材料与方法于2006年9月26日—10月6日分别采集8株叶籽银杏和1株正常银杏(表 1)微黄的种实(胚珠)。采后将叶生种实和正常种实分别标记,种实经脱皮—水冲—阴干—取仁后,连续测定种实、种核和种仁的定性与定量指标。每个指标测30粒,重复3次,数据分析采用SAS软件进行。方差分析采用Yij=μ+τi+ρj+εij随机模型。多重比较采用新复极差检验(Duncan's multiple-range test),即最短显著差数法(SSR-Shortest Significant Ranges)进行。
|
|

叶籽银杏种实形态多样性(表 2)体现在:1) 与正常种实相比(图版Ⅰ-1~4),叶生种实具有明显的外种皮、中种皮、内种皮、胚乳、珠领(图版Ⅰ-5~18);珠孔端和合点端极性明显; 形态上具有多样性(图版Ⅰ-5,13);在缺刻叶脉末端着生1~2(15) 个大小不等的胚珠,但通常只有1枚正常发育(图版Ⅰ-6~9)。成熟的种实外种皮纵隐线和叶片将种实分成不均等的2部分(图版Ⅰ-11),叶片与种实的相连程度一般达种实的1/4~1/3,无种柄。种实上具深褐色的片状、丝状或带状的附属物(图版Ⅰ-10)。种实较正常银杏种实小,呈不规则的心形。具珠领,呈点状或块状,因胚珠嵌入叶片中故均不连续(图版Ⅰ-7~12)。种实顶端渐尖,有的弯曲成喙状(图版Ⅰ-12)。2) 中种皮为骨质硬壳,基端或平或尖,具多束维管束迹(图版Ⅰ-13,14),木质较厚,纹理交错,质地粗糙。种核中上部侧隐线明显或圆润,具二棱或为明显的翼状(图版Ⅰ-15),无麻点或稀麻点。横隐线不明显。珠孔端锐尖,或直立,或为喙状(图版Ⅰ-14),质地均匀,平滑,但较薄。种核较小,为白色或象牙白。种核形状不一,为不规则的心形,背腹稍明显,长宽比较大。3) 内种皮中缢线明显移向珠孔端,上端赤褐色的内种皮缺如或萎缩(图版Ⅰ-16,17)。下端灰白色具蜡层的内种皮明显较正常内种皮扩展,几乎占据了内种皮的全部。4) 剥去内种皮后,可以看见嫩黄色的种仁。种仁外晕为乳白色,内部为浅绿色,未见胚发育仅具胚乳(图版Ⅰ-18),大多胚乳中心有凹陷(或空腔)(图版Ⅰ-18); 种仁圆润,在珠孔端没有明显的突起。
|
|


|
图版Ⅰ Plate Ⅰ 1-4.正常种实; 5-18.叶生种实。1.种实; 2.种核; 3.内种皮及种仁; 4.胚乳与胚; 5.不同大小的叶生种实; 6.一柄2种; 7.叶顶生种实; 8.叶背面着生的种实; 9.近轴面种实退化; 10.外种皮上的附属物; 11.叶生种实的背部和腹部; 12.叶生种实的顶端和基端(珠领); 13.种核的多样性; 14.种核的顶端喙状和基部的多维管束迹; 15. “蚌”状翼; 16.内种皮中线上移; 17.中种皮与内种皮; 18.种仁及胚乳(无胚)。 Aae:远轴端; Ao:败育胚珠; Ap:附属物; Be:基端; Bl:喙状; Ca:空腔; Ce:合点端; Co:珠领; Ds:背面; En:内种皮; Em:胚乳; Eo:胚; Es:叶生种实; Fl:侧棱; L:叶子; Lp:叶柄; Mi:珠孔; Ml:中隐线; Mw:“蚌”状翼; Pae:近轴端; Pe:种托; Pod:顶端; Sa:外种皮; Sc:中种皮; Ss:种柄; Vbt:维管束迹; Vs:腹面。 1-4. Normal seed; 5-18. Epiphyllous seed. 1. Seed; 2. Stone; 3. Endotesta and kernel; 4. Endosperm and embryo; 5. Different size epiphyllous seed; 6. Two epiphyllous seeds on one leaf petiole; 7. Epiphyllous seed on leaf top; 8. Epiphyllous seed on leaf dorsal side; 9. Seed degradation on paraxial end; 10. Appendage on sarcotesta; 11. Dorsal-ventral on epiphyllous seed; 12. Pointed end and basal end on epiphyllous seed; 13. Diversity on stone; 14. Beak-like at pointed end and many vascular bundle traces at basal end; 15. Mussels-wing; 16. Upper shift middle line on endotesta; 17. Sclerotesta and endotesta; 18. Kernel and endosperm(no embryo). Aae: Abaxial end; Ao: Aborted ovule; Ap: Appendage; Be: Basal end; Bl: Beak-like; Ca: Cavity; Ce: Chalazal end; Co: Collar; Ds: Dorsal side; En: Endotesta; Em: Endosperm; Eo: Embryo; Es: Epiphyllous seed; Fl: Flanking; L: Leaf; Lp: Leaf petiole; Mi: Micropylar; Ml: Middle line; Mw: Mussels-wing; Pae: Paraxial end; Pe: Pedicel; Pod: Pointed end; Sa: Sarcotesta; Sc: Sclerotesta; Ss: Seed stalk; Vbt: Vascular bundle trace; Vs: Ventral side. |
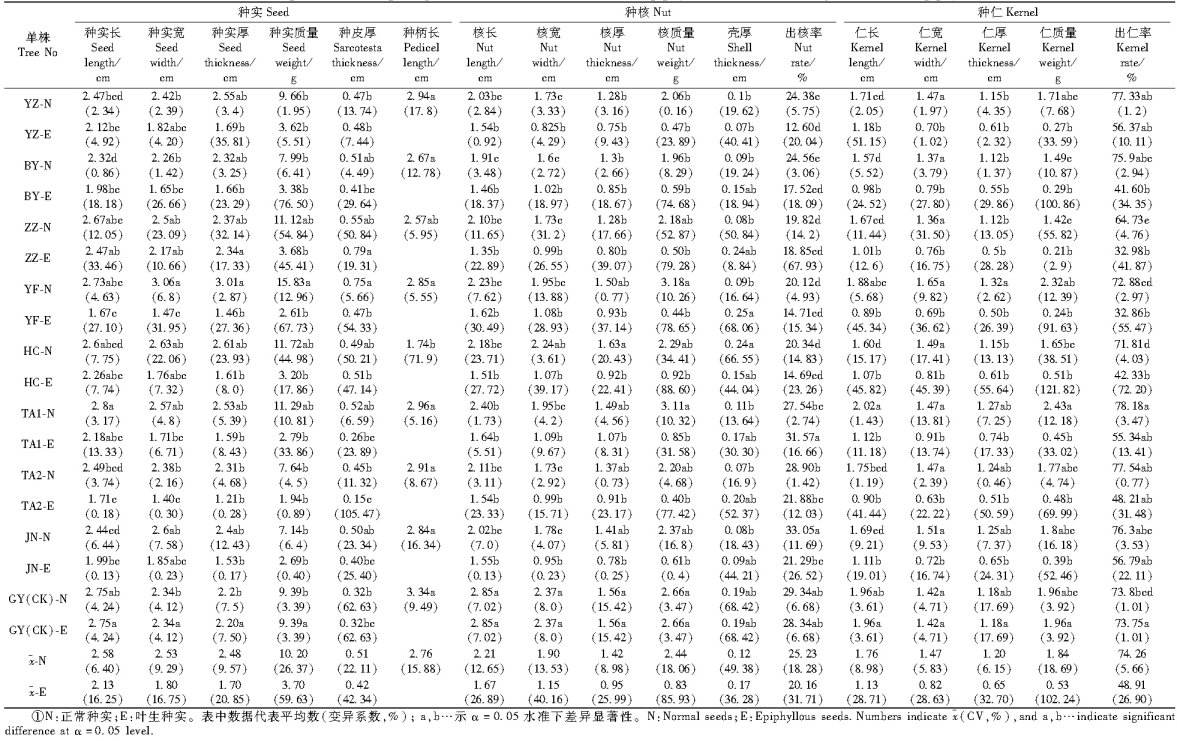
方差分析(表 3)表明:1) 从种实性状来看,正常种实长为2.58 cm,叶生种实为2.13 cm,前者为后者的1.21倍; 种实宽分别为2.53 cm,1.80 cm和1.4倍; 种实厚分别为2.48 cm,1.70 cm和1.46倍; 种实质量分别为10.20 g,3.70 g和2.76倍; 种实皮厚分别为0.51 cm,0.42 cm和1.21倍。2) 从种核性状来看,正常种核和叶生种核的长分别为2.21 cm,1.67 cm,前者为后者的1.32倍; 核宽分别为1.90 cm,1.15 cm和1.65倍; 核厚分别为1.42 cm,0.95 cm和1.50倍; 核质量分别为2.44 g,0.83 g和2.94倍; 壳厚分别为0.12 cm,0.17 cm和0.71倍;出核率分别为25.23%,20.16%和1.25倍。3) 从种仁性状来看,正常种仁和叶生种仁的长分别为1.76 cm,1.13 cm,前者为后者的1.56倍; 仁宽分别为1.47 cm,0.82 cm和1.79;仁厚分别为1.20 cm,0.65 cm和1.85倍; 种仁质量分别为1.84 g,0.53 g和3.47倍; 出仁率分别为74.26%,48.91%和1.52倍。可见正常种实较叶生种实明显大而饱满,出核率、出仁率高。
|
|
本文将含有肉质外种皮的种子称“种实”,去掉外种皮的种子称“种核”,去掉中种皮的种子称“种仁”(邢世岩等,1993)。
Foster等(1974)认为,胚珠是银杏的雌性生殖器官。Zozeran(1949)认为,从短枝上胚珠着生位置来看,银杏雌花是真正的一种原始球果,一个成熟的胚珠由珠心、珠被、珠领—王伏雄等(1983)称珠托组成。银杏大孢子囊产生胚珠,胚珠的珠心经大孢子母细胞→四分体→功能大孢子→雌配子体(邢世岩等,1996; Friedman,1987; 堀輝三,1996)。雌配子体,除产生生殖器官—颈卵器外,其余大部分雌配子体细胞发育成营养组织—胚乳,严格意义上讲胚乳仅是雌配子体的一部分(邢世岩等,1996),颈卵器内产生中央细胞→卵细胞,卵细胞与精子结合形成胚。银杏胚乳是由大孢子母细胞经减数分裂直接发育而成,属于单倍体,而被子植物胚乳为三倍体。银杏内、中、外3层珠被最终发育成膜质的内种皮、骨质中种皮和肉质外种皮。珠领(珠托)发育成种托(或称蒂盘),珠柄发育而成种柄(邢世岩,1993; 邢世岩等,1996; Friedman,1987)(图 1)。

|
图 1 银杏胚珠(A)、正常种实(B)和叶生种实(C)的构造 Figure 1 Structure of ovule(A), normal seed(B) and epiphyllous seed(C) on ginkgo Mi:珠孔; Pc:贮粉室; Po:花粉及花粉管; Nu:珠心; Ar:颈卵器; Fg:雌配子体; In:珠被; Co:珠领; Ch:合点端; Ao:败育胚珠; Os:珠柄; Eo:胚; Em:胚乳; Sa:外种皮; Sc:中种皮; En:内种皮; Pe:种托; Ss:种柄; Ca:空腔; Nes:正常叶生胚珠; Ses:小叶生胚珠; Aes:败育叶生胚珠; L:叶子; Lp:叶柄。 Mi: Micropylar; Pc: Pollen chamber with pollen; Po: Pollen and pollen tubes; Nu: Nucellus; Ar: Archegonium; Fg: Female gametophyte; In: Integuments; Co: Collar; Ch: Chalaza; Ao: Aborted ovule; Os: Ovulate stalk; Eo: Embryo; Em: Endosperm; Sa: Sarcotesta; Sc: Sclerotesta; En: Endotesta; Pe: Pedicel; Ss: Seed stalk; Ca: Cavum; Nes: Normal epiphyllous seed; Ses: Small epiphyllous seed; Aes: Aborted epiphyllous seed; L: Leaf; Lp: Leaf petiole. |
正常种实中种皮和内种皮沿种实纵轴线的赤道面上可见到明显的上下分界线,种体被分成上、下2部分,其形态有别(邢世岩等,1996);但叶生种实,赤道面横隐线明显上移,甚至不发育,种仁几乎被下部的质地脆且不透明、灰白色的内种皮覆盖,这表明叶生种实珠孔端发育不良或退化可能与珠心组织发育异常有关,但仍需继续研究。作者发现,许多叶生种实种核的上部总有空腔发育,而合点端的种仁十分饱满,这从另一方面证明,叶生胚珠珠孔端珠心组织发育异常或退化也许是造成种实无胚的直接原因。
珠被的基部有一边缘突起,即珠领(collar),成熟后称蒂盘。关于银杏珠领的起源及其在裸子植物和种子植物进化中的作用,系统学家和形态学家给予了相当多的关注,关于胚柄的本质不同学者也提出了相关的假说。珠领已被解释为残存的孢子叶,但Pankow等(1967)认为珠领并无维管束,珠领与珠被或外苞叶是不同源的。Florin(1949)认为珠领是一个与胚珠有关的次生性结构(secondary feature),不是叶片。Foster等(1974)认为珠领不具备“叶”的特征,Van Tieghem(1869)认为珠领是残存的假种皮。Douglas等(2007)认为,珠领是银杏特有的构造,在珠被和珠心分化后,靠近胚珠的下皮层细胞进行平周分裂形成珠领。正常种实的种托是连续的,而叶生种实的种托是间断的突起状,围绕在种实的基部,这也许与叶片组织的嵌入有关。对珠领的形态解剖进行研究,对于理解银杏的形态学本质有重要意义。Oda等(1998)认为,银杏种核的壳体以背缝线和腹缝线组成的截面为对称面(法兰面),种缝线在核体的中上部渐趋明显。这与邢世岩等(1993)的结论是一致的。
3.3 叶籽银杏种实形态多样性除了无胚外,成熟的叶生种实的构造与正常种实没有区别,种孔端和合点端极性明显。但种实、种核的形状、大小、顶端、种托具有多样性。种核基部具多点维管束迹,质地粗糙。种核中上部一侧具明显的翼状结构,核体横隐线不明显,顶端为喙状,核体长宽比较大。内种皮中缢线明显移向珠孔端,上端赤褐色的内种皮缺如或萎缩。正常种实和叶生种实质量分别为10.20 g,3.70 g; 正常种核和叶生种核质量分别为2.44 g,0.83 g,出核率分别为25.23%,20.16%;正常种仁和叶生种仁质量分别为1.84 g,0.53 g,出仁率分别为74.26%,48.91%。邢世岩等(1993)证明金坠1# (非叶籽银杏)种实质量11. 84 g,出核率为28.14%,种核质量3.3 g,出仁率81.49%,种仁质量2. 61 g。可见,叶籽银杏正常种实与非叶籽银杏的种实无明显差异,但叶生种实明显质量小,出核率、出仁率低,这也许与胚乳发育不充分有关。
李保进. 2008. 叶籽银杏matK和ITS序列分析及系统发育研究. 山东农业大学硕士学位论文. http://cdmd.cnki.com.cn/Article/CDMD-10434-2008206912.htm
|
李保进, 邢世岩. 2007. 叶籽银杏叶的解剖结构及气孔特性[J]. 林业科学, 43(10): 34-39. DOI:10.3321/j.issn:1001-7488.2007.10.006 |
李士美, 李保进, 邢世岩, 等. 2007. 叶籽银杏拟胚珠的形态发育及变异特性[J]. 园艺学报, 34(1): 1-6. |
福斯特A S, 小吉德福E M. 1983. 维管植物比较形态学. 李正理, 等译. 北京: 科学出版社.
|
王伏雄, 陈祖铿. 1983. 银杏胚胎发育的研究—兼论银杏目的亲缘关系[J]. 植物学报, 25(3): 199-206. |
邢世岩, 高进红, 姜岳忠, 等. 2007. 银杏特异种质核型进化趋势[J]. 林业科学, 43(1): 21-27. |
邢世岩, 皇甫桂月, 侯九寰, 等. 1993. 杏优良单株种子品质的研究Ⅰ.形态品质[J]. 落叶果树, (3): 15-18. |
邢世岩, 孙霞. 1996. 银杏胚胎发育研究述评—兼论银杏系统发育[J]. 武汉植物学研究, 14(3): 279-286. |
邢世岩. 1993. 银杏丰产栽培[M]. 济南: 济南出版社.
|
邢世岩, 樊纪欣, 孙霞, 等. 2005. 银杏的结实特性[J]. 林业科学, 41(1): 30-35. DOI:10.11707/j.1001-7488.20050107 |
邢世岩, 张思清, 张友鹏, 等. 2001. 银杏种子数量遗传分析及多性状选择[J]. 园艺学报, 28(3): 223-229. |
张云跃, 马常耕, 林睦就, 等. 2001. 我国银杏遗传变异研究之一—种核性状的群体间和群体内变异[J]. 林业科学, 37(4): 35-40. DOI:10.11707/j.1001-7488.20010406 |
堀輝三. 1996. ィチョゥの精子—その観察法[J]. 遺传, 50(6): 21-26. |
吉冈金市. 1967. 果树の接木交杂による新种·新品种育成の理论と实际: 第1卷. 东京: 新科学文献刊行会, 143-228.
|
松田孫治. 1932. 秋田縣ニ在ㄦ御葉つきいちょう[J]. 植物研究雜誌, 8(3): 149-450. |
Douglas A W, Stevenson D W, Little D P. 2007. Ovule development in Ginkgo biloba L., with emphasis on the collar and nucellus[J]. Plant Sci, 168(9): 1207-1236. DOI:10.1086/521693 |
Burleigh J G, Mathews S. 2004. Phylogenetic signal in nucleotide data from seed plants:implications for resolving the seed plant tree of life[J]. Am J Bot, 91(10): 1599-1613. DOI:10.3732/ajb.91.10.1599 |
Favre-Duchartre M. 1958. Ginkgo, an oviparous plant[J]. Phytomorphology, 8: 377-390. |
Florin R. 1949. The morphology of Trichopitys heteromorpha Saporta, a seed-plant of Palaeozoic age, and the evolution of the female flowers in the Ginkgoinae[J]. Acta Horti Bergiani, 15: 79-114. |
Foster A S, Gifford E M. 1974. Comparative morphology of vascular plants. 2nd ed.W.H[J]. Freeman and Company. |
Friedman W E. 1987. Growth and development of the male gametophyte of Ginkgo biloba within the ovule(in vivo)[J]. Amer J Bot, 74(12): 1797-1815. DOI:10.2307/2443963 |
Fujii K. 1896. On the different views hitherto proposed regarding the morphology of the flowers of Ginkgo[J]. The Botanical Magazine, 10(108): 7-8. DOI:10.15281/jplantres1887.10.108_7 |
Oda J, Liu Z M, Sintaku S. 1998. Strctural and material composition of a Ginkgo nut shell and its mechanical evaluations[J]. Transactions of the Japan Society of Mechanical Engineers A, 64(624): 2217-2222. DOI:10.1299/kikaia.64.2217 |
Pankow H, Sothmann E. 1967. Histogenetische untersuchungen an den weiblichen blǔten von Ginkgo biloba L[J]. Ber Dtsch Bot Ges, 80: 265-272. |
Sakisaka M. 1927. On the morphological significance of seed-bearing leaves of Ginkgo biloba[J]. The Botanical Magazine, 41(483): 273-278. DOI:10.15281/jplantres1887.41.273 |
Shirai M. 1891. Abnormal Ginkgo tree.Bot Mag Tokyo, 5(56):341-342(in Japanese). https://link.springer.com/chapter/10.1007/978-3-662-02279-5_1
|
Soma S. 1999. Development of the female gametophyte in the ovules on the leaf blade of Ginkgo biloba[J]. Annual Report of the Faculty of Education, Bunkyo University, 33: 112-117. |
Van Tieghem P. 1869. Anatomie comparée de la fleur femelle et du fruit des Cycadées, des Conifères et des Gnétacées[J]. Ann Sci Nat Bot Biol Veg Ser V, 10: 269-304. |
Zozeran R. 1949. Les rameaux courts reproducteure de Ginkgo biloba L[J]. Ann Univ Montpellier Suppl Sci Ser Bot, 4: 68-75. |