 2011, Vol. 47
2011, Vol. 47文章信息
- 张颖娟, 王玉山, 李绍舜
- Zhang Yingjuan, Wang Yushan, Li Shaoshun
- 贮藏条件和时间对西鄂尔多斯5种荒漠植物种子萌发的影响
- Effects of Different Storage Conditions and Durations on Seed Viability and Germination of Five Desert Plants in West Erdos
- 林业科学, 2011, 47(1): 36-41.
- Scientia Silvae Sinicae, 2011, 47(1): 36-41.
-
文章历史
- 收稿日期:2009-04-14
- 修回日期:2010-10-08
-
作者相关文章


2. 内蒙古海拉尔第一中学 海拉尔 021000
2. Haila'er No.1 Middle School Haila'er 021000
种子萌发是植物生命周期的关键阶段,也是植物适应环境变化、保持自身繁衍的重要过程。种子萌发影响到幼苗存活、个体适合度和竞争,进而影响到植被分布和恢复等生态过程(Gutterman, 2000; Donohue et al., 2005)。在我国西北干旱荒漠区,荒漠生态系统是典型而脆弱的系统,在荒漠生态系统对环境变化的响应过程中,种子萌发是关键环节。种子成熟后所经历的环境条件和种子采集后的贮藏条件对种子的萌发行为有显著影响(Meyer et al., 1994; Baskin et al., 1998; 路宁娜等,2008;王磊等,2008;王桔红等,2009)。因此,研究种子贮藏条件、影响种子萌发的环境因子及植物的更新生态位,对干旱区荒漠植被的保护和重建将起重要作用。
荒漠植物种子一般在夏末、秋季成熟,如果种子成熟后立即在这个季节萌发,幼苗往往不能度过寒冷的冬季。种子为了完成生活史过程,必须贮藏到来年春天再萌发。此外,许多植物的种子有休眠的特性,即须经过一段时间的贮藏后才可打破休眠。因此,贮藏持续的时间和条件会影响种子生活力和萌发力,但不同物种的反应可能存在差异(Schütz et al., 1999; van der Valk et al., 1999)。
超旱生灌木和半灌木植物霸王(Zygophyllum xanthoxylum)、沙冬青(Ammopiptanthus mongolicus)、四合木(Tetraena mongolica)、长叶红砂(Reaumuria trigyna)及蒙古扁桃(Amygdalus mongolica)是西鄂尔多斯的优势物种。其中除霸王外,其他物种为古老的珍稀植物,它们的种群维持对稳定荒漠植被和植被恢复具有重要作用。虽然关于这些物种种子萌发特性已有报道(丁琼等, 2006; 李毅等, 2008; 斯琴巴特尔等, 2002; 吴素琴等, 1994; 薛焱等, 2007; 曾彦军等, 2004;2005;张颖娟等,2008a;2008b),但贮藏条件和时间对旱生植物种子萌发的影响目前还鲜见报道,相关内容在湿地植物报道较多(Baskin et al., 1989; van der Valk et al., 1999;刘贵华等,2005)。本文研究贮藏条件和时间对这5种植物种子生活力及萌发的影响,探讨荒漠植物的萌发对策,为荒漠地区的生态恢复和重建提供理论依据。
1 材料与方法 1.1 研究区域概况西鄂尔多斯为内蒙古西部荒漠化草原-草原化荒漠的生态环境脆弱带,是内蒙古高原特有属和亚洲中部(中亚东部)植物特有属的分布中心。该地区自然条件恶劣,风大沙多,土壤贫瘠,年降雨量为80~150 mm, 蒸发量为降雨量的20倍以上。光热条件较好,≥10 ℃年积温在3 000~3 400 ℃。植被以多年生灌木和小灌木为主,物种组成贫乏,以霸王、沙冬青、四合木等古老残遗物种为主要建群种。近年来随着人口增加,资源开发加速以及畜牧业迅速发展,植被破坏日益严重(中国科学院自然资源综合考察委员会,1985)。
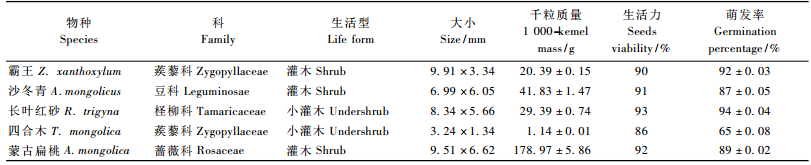
1.2 样品采集与处理试验所用种子来源于各物种集中分布的西鄂尔多斯地区,于2006年各物种成熟的7—9月采集。种子在其自然脱落时人工收集,所采集的种子清理之后,装在信封中备用。每物种选取饱满成熟种子,由于所研究物种的种子在夏秋成熟,经过冬季在翌年春末夏初萌发,考虑种子可能经历的环境条件,设计以下4种贮藏方法:4 ℃冷藏、20 ℃保存、-20 ℃冷冻及野外土壤埋藏CK(种子放在花盆中,上面覆土,埋在试验地)。在贮藏3,6和12个月后,分别测定不同温度贮藏下的种子生活力和萌发率。
1.3 试验方法各物种种子随机抽取100粒1组,用1/10 000天平进行称量,5次重复,求出种子千粒质量及标准差。种子生活力用0.5% TTC染色法测定,具体方法参见张志良等(2003)。
种子萌发率测定时将种子置于培养皿中浸湿的滤纸上,每个培养皿中放置40粒同种植物的种子,每个处理5次重复,在12 h光照1 100 μmol·m-2s-1) :12 h黑暗的光照条件和20 ℃温度条件的光照培养箱中进行萌发,直至连续5天没有种子萌发,结束试验。每天加水维持滤纸湿润,统计种子萌发数。
1.4 统计方法试验数据SPSS 13.0软件进行方差分析,Excel作图。
2 结果与分析 2.1 种子一般特性种子大小、千粒质量表现为种间差异大,种内差异小(表 1)。蒙古扁桃种子最大,千粒质量近179 g,四合木种子千粒质量最小为1.14 g。新采集的成熟完整种子其生活力均在85%以上,具有较高的生活力,并且物种间差异不明显。实验室条件下,除四合木外,其余种子萌发率达85%以上。
|
|
贮藏不同时间后,长叶红砂和蒙古扁桃的种子生活力没有显著变化。长叶红砂低温冷冻(-20 ℃)、蒙古扁桃低温冷藏(4 ℃)时种子生活力最高,但贮藏条件对种子生活力的影响不显著(图 1C,E)。

|
图 1 5种植物在不同贮藏条件和时间的种子生活力 Figure 1 Seed viability of five species at various storage conditions and times A.霸王Z. xanthoxylum; B.沙冬青A.mongolicus;C.长叶红砂R.trigyna;D.四合木T.mongolica;E.蒙古扁桃A.mongolica.下同。The same below. |
霸王、沙冬青和四合木种子生活力随着贮藏时间的增加而逐渐下降。3种植物种子在贮藏3个月时生活力高,贮藏12个月时最低。霸王种子在低温冷冻(-20 ℃)生活力最高,其次是室温下贮藏;沙冬青种子在4 ℃和室温20 ℃生活力较高,而四合木种子在低温冷冻(-20 ℃)和室温贮藏时较高。3种植物种子在野外土壤埋藏时生活力都较低(图 1A,B,D)。
2.3 不同贮藏条件和时间对种子萌发率的影响5种植物种子萌发率的变化如图 2。贮藏3个月时,种子萌发率最高,种子萌发率随着贮藏时间的增加而总体下降,但不同物种在不同贮藏条件下略有差异。

|
图 2 5种植物种子在不同贮藏条件和时间的萌发率 Figure 2 Seed germination percentage of five species at various storage condition and time |
贮藏3个月后,霸王种子萌发率在60%以上,但12个月后萌发率迅速降低,4 ℃贮藏的萌发率不足20%。其中,在室温20 ℃贮藏条件下的萌发率高于其他贮藏条件,4 ℃贮藏条件下,萌发率随贮藏时间增加下降的程度最大。通过两因素方差分析(表 2)表明:不同贮藏条件下,霸王种子萌发率的差异不显著(P=0.063),而不同贮藏时间下萌发率差异显著(P=0.005)。
|
|
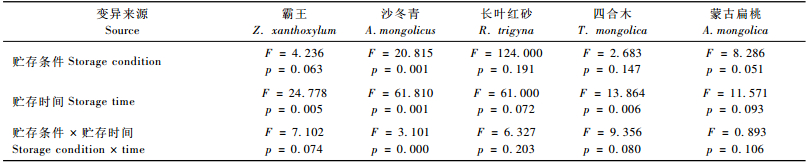
沙冬青种子保存3个月的萌发率在80%以上,6个月后最低的萌发率为63%(野外埋藏)。其中,野外土壤埋藏的种子萌发率明显低于室温20 ℃和4 ℃保存的。两因素方差分析(表 2)表明:贮藏条件和时间对沙冬青种子的萌发率影响极显著(P=0.001)。
贮藏条件和时间对长叶红砂和蒙古扁桃种子的萌发率影响不显著(表 2),但-20 ℃贮藏时种子萌发率均最高,贮藏12个月后长叶红砂种子萌发率仍在90%以上。蒙古扁桃种子在不同贮藏条件下的萌发率均在85%以上。贮藏条件对四合木种子的萌发率影响不显著(P=0.147),但贮存时间对萌发率影响极显著(P=0.006)(表 2)。-20 ℃贮藏时四合木种子萌发率最高,贮藏3个月后萌发率最高为51%,12个月后为18%。
3 结论与讨论 3.1 贮藏条件和时间对种子活力的影响在植物生活周期中,种子对外界极端环境的抵抗力最强,种子对环境的适应能力是植物对环境适应性的重要体现。而幼苗却极易受到不利环境条件的影响,并死亡率较高(Gutterman,2000)。许多学者对特殊生境中种子生态学进行了研究(黄振英,2003;黄振英等,2000a;2000b;聂春雷等,2005;Baskin et al., 1998; Gutterman, 2000)。
每种荒漠植物都具有复杂的生态适应机制,以确保在恶劣的荒漠环境中生存和发展。较多的荒漠植物种子不存在休眠,但种子成熟后的环境条件影响萌发(Baskin et al., 1998),多数物种要到翌年春季以后萌发,贮藏时间影响种子生活力和发芽率。对多数物种而言,随着贮藏时间的延长,种子生活力和发芽率一般都会下降,例如室温24 ℃贮藏超过6个月的苔草(Carex spp.)种子不能萌发( van der Valk et al., 1999)。但不同的物种对贮藏时间的反应会有一定的差异,如一些牧草种子室温25 ℃贮藏几年都能萌发(杜利霞等, 2005),莲(Nelumbo nucifera)的种子在地下深埋几十年仍能保持活力(李曜东等, 2000)。
本试验所选的5种荒漠植物,虽然新采集的种子其生活力均在90%以上(表 1),物种之间差异不显著,但随着贮藏条件和时间的变化,各物种种子生活力的变化不同。
在4种贮藏条件下保存12个月后,长叶红砂和蒙古扁桃的种子生活力仍高达85%~90%。但低温冷冻(-20 ℃)有利于长叶红砂种子保持高生活力;而低温冷藏(4 ℃)则有利于蒙古扁桃种子高生活力的保持。种子保持高生活力可能与种子结构有关,长叶红砂的种子密被绒毛,蒙古扁桃种子具内果皮,均有助于种子的保存,使其保持高生活力。
贮藏时间对霸王、沙冬青和四合木种子生活力的变化影响明显,保存时间越长,种子生活力越低。霸王、沙冬青及四合木贮藏12个月的种子生活力明显低于贮藏3个月的。种子在3个月的短期贮藏,可保持90%左右的生活力。-20 ℃的低温适合霸王种子的贮藏,20 ℃室温相对较适合沙冬青种子的贮藏,四合木种子只适宜短期内保存。因此,即使有大量的种子进入土壤库,在荒漠严酷的自然环境下,种子生活力的降低严重影响着未来的萌发,将导致种群自然更新困难,不利于植被恢复。
3.2 储存条件和时间对种子萌发率的影响贮藏条件和时间对长叶红砂和蒙古扁桃种子萌发率的影响不显著,保存12个月萌发率仍可保持近90%。高生活力和高萌发率虽然有利于种群的扩大和更新,但萌发后一旦遭遇不利的气候变化,很可能导致幼苗大批死亡。野外实际调查也发现,100 m2长叶红砂样地中的幼苗平均仅3~4株。在不可预测的荒漠环境中,种子具有的这种萌发特性可能成为其濒危的一个重要原因。
霸王、沙冬青和四合木的种子萌发率都随着贮藏时间而下降,贮藏3个月的萌发率明显高于12个月的,且差异显著(表 2)。珍稀植物沙冬青的种子萌发率虽然在降低,但野外土壤埋藏保存12个月的种子萌发率仍接近60%,推测沙冬青种子的萌发率在较长时间内可维持在50%以上,有利于土壤中的种子在不同时间萌发,分摊风险,避免一次性萌发导致大批幼苗死亡。濒危种四合木的种子在野外土壤埋藏保存3个月萌发率只有30%,12个月后仅为10%。因此,土壤中的四合木种子经过漫长寒冷的冬季后,即使来年春季条件适宜,能萌发的种子也很少,不利于种群的更新。种子不耐贮藏和萌发率降低可能是四合木濒危的一个原因。
结果表明, 短期贮藏的长叶红砂和蒙古扁桃种子能保持高萌发率, 可作为恢复荒漠植被的物种,但应注意播种季节,选择气候变化小的春末夏初,尽量避免气候突变导致的大批幼苗死亡。霸王、沙冬青和四合木经短期贮藏种子萌发率降低,须对种子进行处理,选择较好保存条件,才能作为植被恢复的物种资源。
丁琼, 王华, 贾桂霞, 等. 2006. 沙冬青种子萌发及幼苗生长特性[J]. 植物生态学报, 30(4): 633-639. |
杜利霞, 李青丰, 刘义. 2005. 不同贮藏时间对牧草种子萌发特性的影响[J]. 中国草地, 27(1): 17-21. |
黄振英, Gutterman Y, 胡正海, 等. 2000a. 沙漠植物种子的传播和萌发机制//李承嘉. 植物科学进展, 第三卷. 北京: 高等教育出版社, 169-178.
|
黄振英. 2003. 鄂尔多斯高原固沙禾草沙鞭种子休眠和萌发与环境的关系[J]. 西北植物学报, 23(7): 1128-1133. |
黄振英, GuttermanY. 2000b. 油蒿与中国和以色列沙漠中的两种蒿属植物种子萌发策略的比较[J]. 植物学报, 42(1): 71-80. |
李曜东, 魏玉凝, 徐本美. 2000. 古莲子与现代莲子ABA含量和SOD活性的比较研究[J]. 植物学通报, 17(5): 439-442. |
李毅, 屈建军, 安黎哲. 2008. 霸王种子萌发的生理条件[J]. 植物生理学通讯, 44(2): 276-278. |
刘贵华, 袁龙义, 苏睿丽, 等. 2005. 贮藏条件和时间对六种多年生湿地植物种子萌发的影响[J]. 生态学报, 25(2): 371-374. |
路宁娜, 崔现亮, 王桔红, 等. 2008. 不同贮藏条件和光照对5种蒺藜科植物种子萌发的影响[J]. 中国沙漠, 28(6): 1130-1135. |
聂春雷, 郑元润. 2005. 鄂尔多斯高原4种主要沙生植物种子萌发与出苗对水分和沙埋的响应[J]. 植物生态学报, 29(1): 32-41. |
斯琴巴特尔, 满良. 2002. 蒙古扁桃种子萌发生理研究[J]. 广西植物, 22(6): 564-566. |
王磊, 严成, 魏岩, 等. 2008. 温度、盐分和储藏时间对多花柽柳种子萌发的影响[J]. 干旱区研究, 25(6): 797-801. |
王桔红, 张勇. 2009. 贮藏条件和温度对4种蒺藜科植物种子萌发的影响[J]. 草业科学, 26(6): 110-115. |
吴素琴, 李克昌, 杨瑞全, 等. 1994. 四合木种子特性的测定研究[J]. 草业科学, 11(3): 29-31. |
薛焱, 王迎春. 2007. 光照、温度和盐分对长叶红沙种子萌发的影响[J]. 植物生理学通讯, 43(4): 708-710. |
曾彦军, 王彦荣, 保平, 等. 2005. 几种生态因子对红砂和霸王种子萌发与幼苗生长的影响[J]. 草业学报, 14(5): 24-31. |
曾彦军, 王彦荣, 庄光辉, 等. 2004. 红砂和霸王种子萌发对干旱与播深条件的响应[J]. 生态学报, 24(8): 1629-1634. |
张颖娟, 李青丰. 2008a. 长叶红砂和红砂种子萌发对光照、温度和水分的响应[J]. 干旱区资源与环境, 22(10): 167-171. |
张颖娟, 王玉山, 李青丰. 2008b. 濒危小灌木长叶红砂种子特性研究[J]. 应用生态学报, 19(12): 2572-2576. |
张志良, 瞿伟菁. 2003. 植物生理学实验指导[M]. 北京: 高等教育出版社: 258-259.
|
中国科学院自然资源综合考察委员会. 1985. 内蒙古植被[M]. 北京: 科学出版社.
|
Baskin C C, Baskin J M, Chester E W. 1989. Role of temperature, light and date: seeds were exhumed from soil on germination of four wetland perennials[J]. Aquat Botany, 35(3/4): 387-394. |
Baskin C C, Baskin J M. 1998. Seeds: ecology, biogeography, and, evolution of dormancy and germination[J]. San Diego: Academic Press: 18-60. |
Donohue K, Dorn L, Griffith C, et al. 2005. The evolutionary ecology of seed germination of Arabidopsis thaliana: variable natural selection on germination timing[J]. Evolution, 59(4): 758-770. DOI:10.1111/evo.2005.59.issue-4 |
Gutterman Y. 2000. Environmental factors and survival strategies of annual plant species in the Negev Desert, Israel[J]. Plant Species Biology, 15(2): 113-125. DOI:10.1111/psb.2000.15.issue-2 |
Meyer S E, Kitchen S G. 1994. Habitat correlated variation in seed germination response to chilling in Penstemon Section Glabri (Scrophulariaceae)[J]. American Midland Naturalist, 132(2): 349-365. DOI:10.2307/2426591 |
Schütz W, Rave G. 1999. The effects of cold stratification and light on the seed germination of temperate sedges (Carex) from various habitats and implications for regenerative strategies[J]. Plant Ecology, 144(1): 215-230. |
van der Valk A G, Bremholm T L, Gordon E. 1999. The restoration of sedge meadows: seed viability, seed germination requirements, and seedling growth of Carex species[J]. Wetlands, 19(4): 756-764. DOI:10.1007/BF03161782 |