 2011, Vol. 47
2011, Vol. 47文章信息
- 张象君, 王庆成, 王石磊, 孙强
- Zhang Xiangjun, Wang Qingcheng, Wang Shilei, Sun Qiang
- 小兴安岭落叶松人工纯林近自然化改造对林下植物多样性的影响
- Effect of the Close-to-Nature Transformation of Larix gmelinii Pure Stands on Plant Diversity of Understory Vegetation in Xiaoxing'an Mountains of China
- 林业科学, 2011, 47(1): 6-14.
- Scientia Silvae Sinicae, 2011, 47(1): 6-14.
-
文章历史
- 收稿日期:2009-11-12
- 修回日期:2010-10-07
-
作者相关文章



人工林(尤其是针叶纯林)在其培育过程中会出现生物多样性严重下降(Paritsis et al., 2008; Tiefenbacher, 1999; 张文臣,2004; 杨学云,2005)、林分生态功能低下和生产力下降(Gamborg et al., 2003; 陆元昌,2006)等一系列问题。目前,根据分类经营的要求,一些人工纯林被划入生态公益林范畴,如何经营这部分人工纯林,提高其生态功能和生产力就显得尤为重要。
人工林近自然化改造是以天然森林的结构和演替过程为参照,模拟自然进程来改造人工纯林,使其近似于天然森林的复层异龄结构,从而提高生物多样性和生产力水平(陆元昌,2006)。目前,人工林近自然化改造的主要措施为目标树单株择伐,需要经过多次作业才能完成(Abetz et al., 2002; 李慧卿等,2007; 陆元昌,2006; 束传林,2007; 郝云庆等,2005)。根据目标树种生长能力的不同,在北方地区目标树单株择伐改造时间一般为40~60年,甚至更长(陆元昌,2006)。而东北地区主要人工林—落叶松(Larix gmelinii)人工林在40~60年为成熟林,60年后即达到过熟状态(卢伟,2003),如果采用以上方式改造,很可能会由于同龄主林层的迅速老化带来生长退化,甚至主林层消失,导致改造失败。近年来,林隙干扰研究已成为森林生态学和森林经营学研究最活跃的领域之一,林隙的形成和变化构成了森林景观的流动镶嵌结构,对林隙的研究揭示了森林动态变化机制,为森林可持续经营及生物多样性维护提供了必要的科学依据(田悦颖,2007)。在此基础上,一些学者提出采用林隙间伐(或群团状采伐)的方式对人工林进行近自然化改造的设想,并进行了相关试验研究,结果表明:林隙间伐通过模拟小尺度自然干扰形成的林隙来增加生境异质性和林分结构的复杂性,从而缩短了近自然化改造的进程(Macolm et al., 2001; Mason, 2003; Albanesi et al., 2005; Hiromi, 2007; Davis et al., 2007; Dekker, 2008; Fahey et al., 2008)。
目前,国内外众多学者对林隙内植物多样性的研究大多表明林隙促进了植被发育、提高了植物多样性,不同大小林隙对林下植被的影响差异较大(Fahey et al., 2008; Schnitzer et al., 2001; 钱莲文等, 2005; 齐代华等,2006; 南海龙等,2006; 臧润国等,2000),但大多只局限于林隙内植被的变化(Fahey et al., 2008; 王周平等,2003; 张艳华等,1999; 龙翠玲,2009),笔者认为研究人工林近自然化改造(林隙间伐)对植物多样性的影响应该着眼于林分水平,而非仅局限于林隙水平。常规均匀间伐是目前人工林经营的主要方式,在促进林下植被发育等诸多方面发挥了重要作用(徐扬等,2008; 李春义等,2006),但与林隙间伐相比,二者对林下植被的影响是否存在差异?林隙大小虽对林下植被影响较大,但在林分水平上,相同间伐强度的不同大小林隙间伐方式对林下植被产生的影响是否会存在较大差异?凋落物覆盖对林下植被影响比较复杂(高志红等,2004; 段昌群,2006), 通过在林隙内整地是否能显著改变林下植被?本研究对黑龙江省主要针叶人工林—落叶松人工林进行不同大小林隙间伐,并辅以整地进行近自然化改造,以相同强度的常规均匀间伐处理为对照,揭示近自然化改造和常规均匀间伐对林下植物多样性的影响,为以林隙间伐为基础的人工林近自然化改造提供理论依据。
1 研究区概况研究在黑龙江省带岭林业局东方红林场进行。林场位于黑龙江省东北部、小兴安岭南坡(128°57′16″—129°17′50″E,46°50′8″—46°59′20″N)。该区以低山为主,海拔250~1 050 m。温带大陆性季风气候,冬季漫长,寒冷干燥,夏季短促,炎热。年平均气温1.4 ℃,1月平均气温-19.4 ℃,极端最低气温-40 ℃,7月平均气温20.9 ℃,极端最高气温37 ℃。大于10 ℃年积温2 156 ℃,全年无霜期115天。年平均降水量661 mm,主要集中于7—9月份,占全年降水量的50%以上。地带性土壤为暗棕壤。
研究林分位于平缓山地,平均坡度为8°,立地条件变化较小。林分的树种组成为18年生落叶松,混有少量山桃稠李(Padus maackii)和白桦(Betula platyphylla)(比例小于2%),林木分布均匀,平均胸径13.4 cm,平均树高11.6 m,林分密度3 704株·hm-2。林下主要植物有东北山梅花(Philadelphus schernkii)、瘤枝卫矛(Evonymus verrucosus)、光萼溲疏(Deutzia glabrata)、黄花忍冬(Lonicera chrysantha)、毛缘苔草(Carex campylorhina)、狭叶荨麻(Urtica angustifolia)、白花碎米荠(Cardamine leucantha)、灰绿藜(Chenopodium glaucum)、乌苏里黄芩(Scutellaria pekinbensis var. ussuriensis)、白屈菜(Chelidonium majus)、深山露珠草(Circaea caulescens)和林地早熟禾(Poa nemoralis)等。
2 研究方法2008-03-01—05-01,在18年生落叶松人工纯林中实施相同株数强度(25%)的林隙间伐,并以相同株数强度的常规均匀间伐处理为对照。林隙采伐处理中,林隙在林分中均匀分布。小林隙、中林隙和大林隙间伐面积分别为25,50和100 m2,林隙形状近似正方形,其边长分别约为5,7和10 m,林隙边长与树高的的比值分别近似为1/2,3/4和1/1。以上4种处理在2个小区中实施即重复2次,小区面积4.0 hm2,每小区每处理均1 hm2,各处理在各小区中随机配置。在第1试验小区的林隙内,带状破草皮并翻垦林隙内的土壤,带宽1.0 m,保留带1.0 m,翻垦深度5 cm(对照区不整地)。第2试验小区的林隙内不做任何处理,保持原状。
分别于2008年7月下旬及2009年7月下旬进行林下植被调查。在各试验小区4种处理内随机设置10个16 m×16 m的大样方。在每个大样方内随机设5个2 m×2 m的灌木样方(可能位于林隙内,也可能位于林隙外),在每个灌木样方内对角位置设2个1 m×1 m的草本样方。记录每一灌木样方内灌木的种数、个体数、树高和盖度,记录草本样方内草本植物的物种数、个体数(丛数)、高度和盖度。将乔木幼苗(高<1 m统计在灌木中。均匀间伐处理的调查仅在第1小区进行。
采用以下指数进行多样性的测定:1) 重要值(Pi) Pi=(Rd+Rde+Rf)/3, Rd为相对密度,Rde为相对优势度,Rf为相对频度; 2) Simpson多样性指数(D) 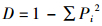
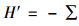
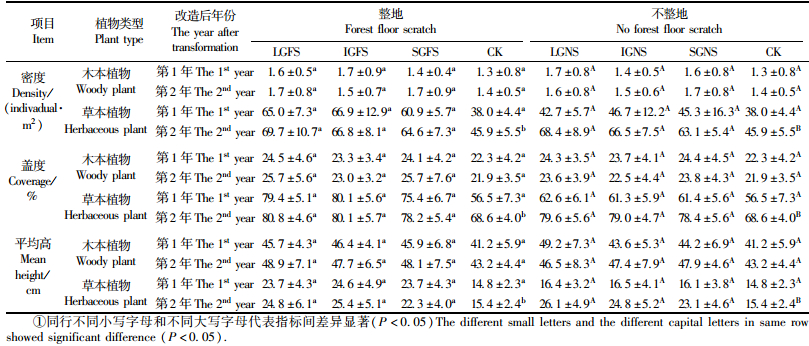
与对照相比,近自然化改造后第1年和第2年林下木本植物的种数虽有所增加,但差异均不显著(P>0.05)(图 1)。改造后第1年,大林隙间伐整地、大林隙间伐不整地、中林隙间伐整地及中林隙间伐不整地林下草本植物种数均显著高于对照(P<0.05),小林隙间伐整地和小林隙间伐不整地虽高于对照,但差异不显著(P>0.05);改造后第2年,各近自然化改造林下草本植物种数均显著高于对照(P<0.05)(图 1)。改造后第1年和第2年,无论整地改造还是不整地改造,林下草本植物种在不同大小林隙间伐改造之间差异不显著(P>0.05),在整地改造与不整地改造之间也无显著差异(P>0.05)(图 1)。

|
图 1 近自然化改造对林下植物种数的影响 Figure 1 Effect of close-to-nature transformation on species number of understory plants |
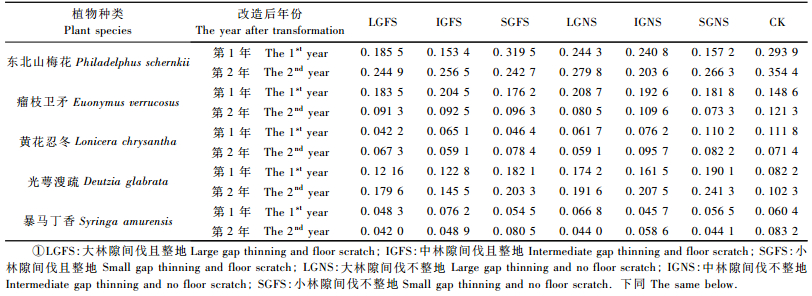
近自然化改造对林下木本植物优势种的影响较小。无论是改造后第1年还是第2年,近自然化改造处理和对照林下木本植物优势种均为东北山梅花、瘤枝卫矛、黄花忍冬、光萼溲疏和暴马丁香(Syringa amurensis)(表 1)。与对照相比,改造后第1年和改造后第2年林下光萼溲疏的重要值明显增加,而瘤枝卫矛、东北山梅花和暴马丁香重要值在改造后第2年降低,黄花忍冬重要值的变化无明显趋势。不同改造处理间,主要木本植物重要值的变化未发现明显规律(表 1)。
|
|
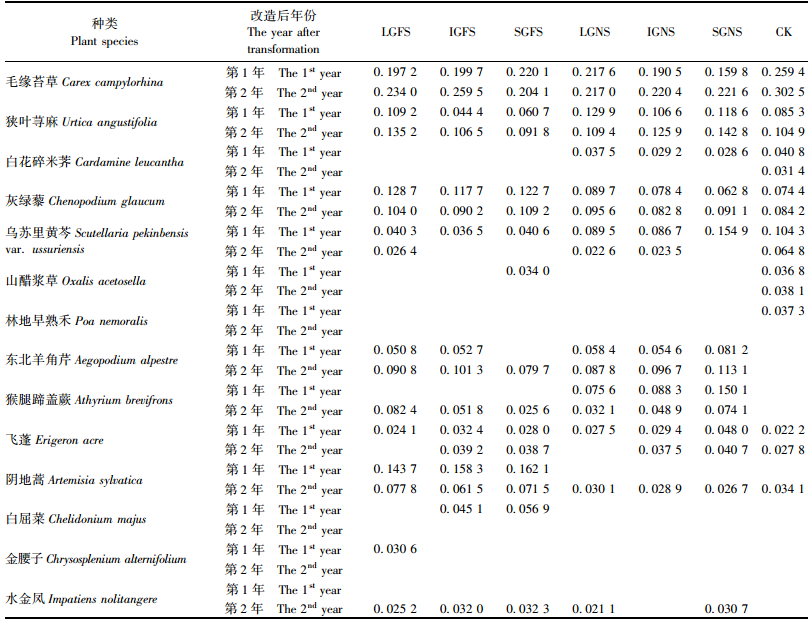
近自然化改造对林下草本植物优势种(重要值排在前8位)的影响较大。与对照相比,改造后第2年近自然化改造的草本植物优势种出现了东北羊角芹(Aegopodium alpestre)、猴腿蹄盖蕨(Athyrium brevifrons)和水金凤(Impatiens nolitangere)等,而对照林下草本植物优势种白花碎米荠、山醋浆草(Oxalis acetosella)和乌苏里黄芩等的重要值在各近自然化改造林分中均明显下降(表 2)。不同大小林隙间伐改造之间(无论是整地还是不整地),林下草本植物优势种的变化无明显差异(表 2)。改造后第1年,与不整地改造相比,无论是大林隙、中林隙还是小林隙,整地改造林下草本植物优势种中均出现了阴地蒿(Artemisia sylvatica),而不整地改造林下的优势种白花碎米荠和猴腿蹄盖蕨则未成为整地改造林下的优势种; 改造后第2年,整地改造和不整地改造之间林下草本植物优势种未发现明显差异(表 2)。
|
|
与对照相比,近自然化改造后第1年和第2年林下木本植物的密度、盖度和平均高均有不同程度的提高,但差异均不显著(P>0.05)(表 3)。与对照相比,改造后第1年林隙间伐辅以整地改造林分的草本植物密度显著增加(P<0.05),而林隙间伐不整地与对照的差异不显著(P>0.05);改造后第2年,各近自然化改造的草本植物密度均显著高于对照(P<0.05),其中,大林隙间伐整地和大林隙间伐不整地的草本植物密度增幅较大,分别比对照增加51.9%和49.0%;小林隙间伐整地和小林隙间伐不整地草本植物密度的增幅较小,分别比对照增加40.7%和37.5%(表 3)。无论是整地改造还是不整地改造,其不同大小林隙间伐之间林下草本植物密度在改造后第1年和第2年差异均不显著(P>0.05)(表 3)。改造后第1年,无论是大林隙、中林隙还是小林隙处理,整地的林下草本植物密度均显著高于不整地的(P<0.05),改造后第2年差异显著性消失(P>0.05)(表 3)。近自然化改造对林下草本植物盖度和平均高的影响与近自然化改造对林下草本植物密度的影响相同。与对照相比,改造后第2年大林隙间伐整地改造林下草本植物盖度增幅最大,增加了17.8%;小林隙间伐不整地增幅较小,为14.3%。林下平均高增幅最大的为大林隙间伐不整地,增加了69.5%;增幅较小的为小林隙间伐整地,增加了44.8%(表 3)。
|
|
与对照相比,各近自然化改造处理在改造后第1年和第2年林下木本植物Simpson指数、Shannon-Wiener指数和均匀度指数均未发现显著的变化(P>0.05)(图 2)。

|
图 2 近自然化改造对林下木本植物多样性的影响 Figure 2 Effect of close-to-nature transformation on the diversity of understory woody plants |
与对照相比,改造后第1年各近自然化改造林下草本植物Simpson指数、Shannon-Wiener指数和均匀度指数均有不同程度的升高,除中林隙间伐不整地的Simpson指数和林隙间伐整地的均匀度指数外,差异不显著(P>0.05)(图 3); 改造后第2年,各近自然化改造的Simpson指数、Shannon-Wiener指数显著高于均匀间伐(P<0.05),均匀度指数虽高于均匀间伐,但差异不显著(大林隙间伐整地和中林隙间伐不整地除外)(P>0.05)(图 3)。改造后第1年和第2年,无论是整地改造还是不整地改造,不同大小林隙改造之间的Simpson指数、Shannon-Wiener指数和均匀度指数差异均不显著(P>0.05);无论是大林隙、中林隙还是小林隙,整地改造和不整地改造之间的Simpson指数、Shannon-Wiener指数和均匀度指数差异均不显著(P>0.05)(图 3)。

|
图 3 近自然化改造对草本植物多样性的影响 Figure 3 Effect of close-to-nature transformation on the diversity of herbaceous plants |
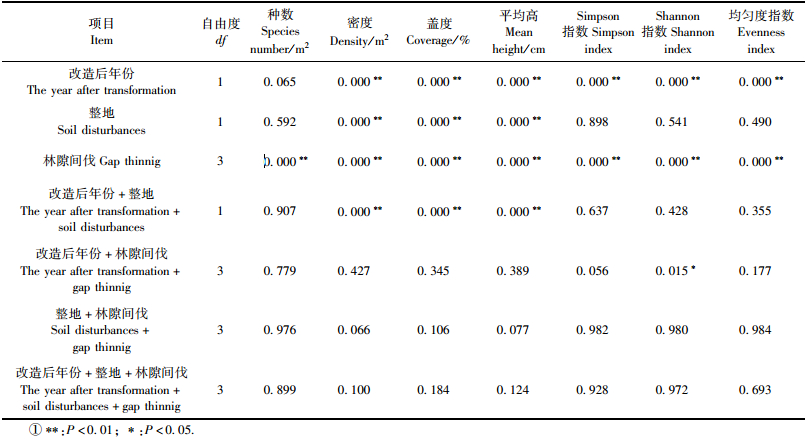
近自然化改造对草本植物影响的方差分析详见表 4。林隙间伐和改造后年份等因素对林下草本植物种数、密度、盖度、平均高和多样性指数均产生影响,除改造后年份对种数影响的差异为P=0.065外,差异均达到极限显著(P<0.01);整地处理只对密度、盖度和平均高产生极显著影响(P<0.01);改造后年份+整地的交互作用也只对密度、盖度和平均高产生极显著影响(P<0.01);改造后年份+林隙间伐的交互作用只对Shannon指数产生显著影响(P=0.015);林隙间伐+整地的交互作用和改造后年份+整地+林隙间伐的交互作用未能对林下草本植物种数、密度、盖度、平均高和多样性指数产生影响(P>0.05)。近自然化改造未能对林下木本植物产生显著影响,方差分析略。
|
|
与对照(常规均匀间伐)相比,近自然化改造后第2年落叶松人工林林下草本植物种类、密度、盖度、高度和多样性指数(Simpson指数、Shannon-Wiener指数)均显著增加(P<0.05)。常规均匀间伐增加了林下光照、降低了林分密度,使林下获得了充分的资源(光照、温度、养分等),促进了林下植被的发育(马履一等,2007; 李春义等,2006; 毛志宏等,2006)。而近自然化改造(林隙间伐)在增加林下资源的同时,又形成了林隙和非林隙的斑块分布格局,具有更大的异质性,为不同植物的定居和繁殖创造了更加有力的条件,进而提高了林下植物多样性(王进欣等,2002; Fahey et al., 2008; Muscolo et al., 2007; Schnitzer et al., 2001; Zhu et al., 2003)。以往的研究虽也表明林隙的形成促进了植物定居,进而增大个体密度,有利于维持林下植物多样性,但往往仅限于林隙范围内,并且是相对于未间伐林分而言(Cason et al., 2008; Schnitzer et al., 2001; 王周平等,2003; 张艳华等,1999; 龙翠玲,2009),未能很好地证明林隙间伐在维持林下植物多样性方面所发挥的重要作用。
与对照相比,各近自然化改造林下草本植物优势种中出现了羊角芹、猴腿蹄盖蕨、水金凤等喜湿植物(表 3)。这是因为近自然化改造形成的林隙,在林隙小范围内消除了上层乔木的蒸腾消耗以及对降水的截流,在一定时期内明显增加了土壤水分,为喜湿植物的出现创造了有力条件(胡建伟等,1999; 朱教君等,2007)。
改造后第1年和第2年,无论整地还是不整地,其不同大小林隙间伐改造之间林下草本植物种数、密度、盖度、平均高和多样性指数均未发现明显差异。表明当间伐强度相同时,不同大小的林隙间伐改造后林隙植被在林分水平上无明显差异。但也有可能是本研究的林隙梯度不大,这有待于进一步研究。此外,有研究表明,林隙导致的植物多样性增加是暂时的,随着林隙的郁闭,最终多样性还会降低(Schnitzer et al., 2001)。本研究中,不同林隙的郁闭时间不同,随着时间推移,不同大小林隙间伐对林下植被的影响是否仍然没有差异,还有待进一步调查。
改造后第1年,无论大林隙、中林隙还是小林隙,其整地改造的草本植物密度、盖度、平均高均显著高于不整地改造(P<0.05)(表 4)。这是因为林隙间伐整地处理在一定程度上移除了林隙内的枯落层,而枯落物的存在增加了种子萌发的机械阻力、降低土壤表面光照,进而抑制了林下植被发育(高志红等,2004)。改造后第2年,林隙间伐整地处理与林隙间伐不整地处理间草本植物差异不显著(P>0.05)。
与对照处理相比,改造后第1年和第2年近自然化改造林下木本植物种数、密度、盖度、平均高和多样性指数均未发现显著的差异(P>0.05),可能是改造时间短的原因。Hubbell等(1999)对巴拿马地区2龄林隙的林下木本植物(胸径1~4 cm)多样性的研究也发现,林隙未能显著增加物种多样性。近自然改造后,林下更新苗木(如1龄、2龄苗木)在改造初期的更新状况如何,随着时间的延长,各近自然化改造与对照的木本植物多样性是否仍无明显差异,其更新动态如何,都有待进一步研究。
与对照(常规均匀间伐)相比,近自然化改造在短期内更有利于林下草本植物的发育和草本植物多样性的提高。对于林下木本植物,各近自然化改造和对照之间未发现显著差异。
段昌群. 2006. 生态科学进展. 2卷. 北京: 高等教育出版社. http://edu.wanfangdata.com.cn/Periodical/Detail/lsdsj201505009
|
高志红, 张万里, 张庆费. 2004. 森林凋落物生态功能研究概况及展望[J]. 东北林业大学学报, 32(6): 79-83. |
郝云庆, 王金锡, 王启和, 等. 2005. 崇州林场不同林分近自然度分析与经营对策研究[J]. 四川林业科技, 26(2): 20-26. |
胡建伟, 朱成秋. 1999. 抚育间伐对森林环境的影响[J]. 东北林业大学学报, 27(3): 65-67. |
李春义, 马履一, 徐昕. 2006. 抚育间伐对森林生物多样性研究进展[J]. 世界林业研究, 19(6): 27-32. |
李慧卿, 江泽平, 雷静品, 等. 2007. 近自然森林经营探讨[J]. 世界林业研究, 20(3): 6-11. |
龙翠玲. 2009. 茂兰喀斯特森林林隙物种多样性的动态规律[J]. 山地学报, 27(3): 278-285. |
卢伟. 2003. 黑龙江省国有林区营林技术系列标准[M]. 哈尔滨: 东北林业大学出版社.
|
陆元昌. 2006. 近自然森林经营的理论与实践[M]. 北京: 科学出版社.
|
马履一, 李春义, 王希群, 等. 2007. 不同强度间伐对北京山区油松生长及其林下植物多样性的影响[J]. 林业科学, 43(5): 1-9. |
毛志宏, 朱教君, 刘足根, 等. 2006. 间伐对落叶松人工林内草本植物多样性及其组成的影响[J]. 生态学杂志, 25(10): 1201-1207. DOI:10.3321/j.issn:1000-4890.2006.10.010 |
南海龙, 韩海荣, 马钦彦, 等. 2006. 太岳山针阔混交林林隙草本和灌木物种多样性研究[J]. 北京林业大学学报, 28(2): 52-56. |
齐代华, 李旭光, 王周平, 等. 2006. 间伐对落叶松人工林内草本植物多样性及其组成的影响[J]. 生态学杂志, 25(10): 1201-1207. DOI:10.3321/j.issn:1000-4890.2006.10.010 |
钱莲文, 郭建宏, 吴承祯, 等. 2005. 长苞铁杉林林隙梯度物种多样性[J]. 东北林业大学学报, 33(5): 53-55. |
束传林. 2007. 云南松林近自然化采伐的改造效果[J]. 陕西林业科技, (2): 14-17. |
田悦颖. 2007. 小兴安岭阔叶红松林林隙特征及其更新研究. 东北林业大学硕士学位论文. http://www.cnki.com.cn/Article/CJFDTOTAL-DBLY201604008.htm
|
王进欣, 张一平. 2002. 林隙微环境异质性及物种的响应[J]. 南京林业大学学报, 26(1): 69-74. |
王周平, 李旭光, 石胜友, 等. 2003. 缙云山森林林隙与非林隙物种多样性比较研究[J]. 应用生态学报, 14(1): 7-10. |
徐扬, 刘勇, 李国雷, 等. 2008. 间伐强度对油松中龄人工林林下植被多样性的影响[J]. 南京林业大学学报, 32(3): 135-138. |
杨学云. 2005. 浅议我国人工林的近自然经营[J]. 中南林业调查规划, 24(1): 7-9. |
臧润国, 王伯荪, 刘静艳. 2000. 南亚热带常绿阔叶林不同大小和发育阶段林隙的树种多样性研究[J]. 应用生态学报, 11(4): 485-488. |
张文臣, 张俊佩. 2004. 人工林存在的问题与对策[J]. 防护林科技, (3): 345-346. |
张艳华, 聂绍荃, 王志华. 1999. 林隙对草本植物的影响[J]. 植物研究, 19(1): 94-99. |
朱教君, 刘世荣. 2007. 森林干扰生态学研究[M]. 北京: 中国林业出版社.
|
Abetz P, Kladtke J. 2002. The target tree management system[J]. Forstwiss Centralbl, 121(2): 73-82. DOI:10.1046/j.1439-0337.2002.00073.x |
Albanesi E, Gugliotta O I, Mercurio I, et al. 2005. Effects of gap size and within-gap position on seedlings establishment in silver fir stands[J]. Jouranl of Biogeosciences and Forestry, 1: 55-59. |
Cason W P, Schnitzer S A. 2008. Tropical forest community ecology//Schnitzer S A, Mascaro J, Cason W P. Treefall gaps and the maintence of plant species diversity in tropical forests. Oxford: Blackwell Publishing, 196-209.
|
Davis L R, Puettmann K J, Tucker G F. 2007. Overstory response to alternative thinning treatments in young Douglas-fir forests of western Oregon[J]. Northwest Science, 81(1): 1-14. DOI:10.3955/0029-344X-81.1.1 |
Dekker M. 2008. Growth patterns, competition and coexistence in gap-phase regeneration under close-to-nature silviculture[J]. Wageningen University, PhD. |
Fahey R T, Puettmann K J. 2008. Patterns in spatial extent of gap influence on understory plant communities[J]. Forest Ecology and Management, 255(7): 2801-2810. DOI:10.1016/j.foreco.2008.01.053 |
Gamborg C, Larsen J B. 2003. 'Back to nature'—a sustainable future for forestry[J]. Forest Ecology and Management, 179(1/3): 559-571. |
Hiromi M. 2007. Do finer gap mosaics provide a wider niche for Quercus gilva in young Japanese ceder plantations than coarser mosaics? Simulation of spatial heterogeneity of light availability and photosynthetic potential[J]. Canadian Journal of Forest Research, 37(9): 1545-1553. DOI:10.1139/X07-124 |
Hubbell S P, Foster R B, O'Brien S T, et al. 1999. Light-gap disturbances, recruitment limitation and tree diversity in a neotropical forest[J]. Scinece, 283(5401): 554-557. DOI:10.1126/science.283.5401.554 |
Macolm D C, Mason W L, Clarke G C. 2001. The transformation of conifer forests in Britain—regeneration, gap size and silvicultural systems[J]. Forest Ecology and Management, 151(1/3): 7-23. |
Mason W L. 2003. Continuous cover forestry: developing close to nature forest management in conifer plantations in upland Britain[J]. Scottish Forestry, 57(3): 141-149. |
Muscolo A, Sidari M, Mercurio R. 2007. Influence of gap size on organic matter decomposition, microbial biomass and nutrient cycle in Calabrian pine(Pinus laricio, Poriet) stands[J]. Forest Ecology and Management, 242(2/3): 412-418. |
Paritsis J, Aizen M A. 2008. Effects of exotic conifer plantations on the biodiversity of understory plants, epigeal beetles and birds in Nothofagus dombeyi forests[J]. Forest Ecology and Management, 255(5/6): 1575-1583. |
Schnitzer S A, Carson W P. 2001. Treefall gaps and the maintenance of species diversity in a tropical forest[J]. Ecology, 82(4): 913-919. DOI:10.1890/0012-9658(2001)082[0913:TGATMO]2.0.CO;2 |
Tiefenbacher E. 1999. Measuring and controlling the degree of naturalness of forest stand[J]. Zeitschrift fur Foretwesen, 150(7): 246-248. |
Zhu J, Matsuzaki T, Lee F, et al. 2003. Effect of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest[J]. Forest Ecology and Management, 182(1/3): 339-354. |