 2011, Vol. 47
2011, Vol. 47文章信息
- 周国模, 刘恩斌, 施拥军, 刘安兴, 周宇峰
- Zhou Guomo, Liu Enbin, Shi Yongjun, Liu Anxing, Zhou Yufeng
- 基于最小尺度的浙江省毛竹生物量精确估算
- Accurate Estimation for Moso Bamboo (Phyllostachys edulis) Biomass in Zhejiang Province Based on the Lowest Scale Technique
- 林业科学, 2011, 47(1): 1-5.
- Scientia Silvae Sinicae, 2011, 47(1): 1-5.
-
文章历史
- 收稿日期:2010-08-08
- 修回日期:2010-09-19
-
作者相关文章


2. 浙江省森林资源监测中心 杭州 310020
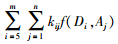 实现区域尺度生物量的精确估算。用该方法估算得浙江省毛竹总生物量为1.471 6×1010 kg。
实现区域尺度生物量的精确估算。用该方法估算得浙江省毛竹总生物量为1.471 6×1010 kg。2. Monitoring Center for Forest Resources, Zhejiang Province Hangzhou 310020
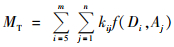 .The estimated results showed that the total moso bamboo biomass in Zhejiang Province was 1.471 6×1010 kg.
.The estimated results showed that the total moso bamboo biomass in Zhejiang Province was 1.471 6×1010 kg. 森林生态系统生物量估算是预估陆地生态系统与大气间温室气体CO2交换量的关键(Dixon et al., 1994), 并成为国际地球生物圈计划(IGBP)、世界气候研究计划(WCRP)和全球环境变化国际人文因素计划(IHDP)等重大科学计划的研究主题。如何准确地对森林生态系统生物量进行估算是近期备受国际科学界广泛关注的重大议题(Brown, 2002)。目前对区域尺度森林生物量的估算主要采取尺度转换的方法(赵敏等,2004;Zhang et al., 2002; Fang et al., 2001),但由于森林生态系统的复杂性以及常用尺度转换方法的缺陷(Brown et al., 1984;Isaev et al., 1995; Johnson et al., 1983;Zhou et al., 2002), 导致不同学者对同一尺度森林植被生物量的估算结果差异较大(Dixon et al., 1994; 周玉荣等, 2000; 方精云等, 1996)。因此,提出一种能直接精确估算区域尺度森林生物量的方法意义重大。毛竹(Phyllostachys edulis)是我国南方的一种特殊植物,具有生长快、周期短、产量高、用途广、固碳功能强(周国模等,2004;周国模,2006)等诸多优点。浙江省毛竹林面积及产量均位居全国前列,本研究以浙江省毛竹为对象,应用二元概率分布函数与单株毛竹生物量模型,以最小尺度单株毛竹(尽管单株毛竹不是生物量计量的最小单位,但对于大区域或大尺度的总生物量而言主,单株生物量是非常小的尺度单位,再考虑到生物量尺度转换的实用性与方便性,本研究把单株毛株看做生物量的最小尺度)为基础,提出一种直接精确估算全省毛竹总生物量的方法,其目的在于:1) 实现定量描述浙江省毛竹的生态功能;2) 为全省毛竹碳交易的国际谈判提供技术支撑;3) 为生物量新尺度转换方法的提出奠定基础。
1 二元Weibull分布函数的推导毛竹是一种特殊的地域性植物,与其他植物不同的是:1) 毛竹的胸径生长与高生长在第1年内完成;2) 对单株毛竹而言,当胸径生长与高生长完成后,胸径与竹高不再发生变化,但其生物量还在变化,生物量的这种变化主要是由年龄引起的;3) 毛竹林是一种异龄林,故除胸径外,年龄也是其林分结构的重要特征。因此影响毛竹生物量的因素为胸径与年龄。
一元Weibull分布由于其灵活性大、适应性强且积分形式简单,在实践中常用来作为测树因子概率分布模型,但它只能描述一个测树因子的概率分布规律,不能测量毛竹胸径和年龄的联合概率分布信息,而二元Weibull分布可以弥补一元分布的这种不足。有关二元Weibull分布的应用研究已有报道(葛宏立等,2008),但在理论上对二元Weibull分布进行严密推导的研究尚未见报道。本研究构建二元Weibull分布模型并用它来测量浙江省毛竹胸径和年龄的联合分布信息。
二元Weibull分布生存函数(史道济等, 2003; Shi, 1995; Tawn,1988)为:
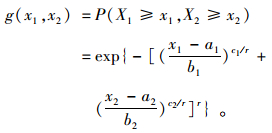
|
(1) |
式中X1和X2为2个随机变量,x1和x2分别为他们的取值;a1和a2为位置参数,b1和b2为尺度参数;c1和c2为形状参数,x1>a1, x2>a2。在区域x1≤a1, x2≤a2内,概率密度定义为0, b1, b2, c1和c2均大于0, r(0 < r≤1) 为相关系数。
从生存函数的表达式可以看出:随着x1, x2增大,g(x1, x2)值减小,故Weibull生存函数能反应植物的生存概率。
现应用二元Weibull生存函数与图 1(G1,G2,G3和G4为积分区域)对二元Weibull分布函数做如下推导。

|
图 1 二元Weibull生存函数与分布函数积分区域示意 Figure 1 Bivariate Weibull survival function and distribution function integration regions |
设x1, x2定义域的下界分别为a1, a2(如果x1, x2是测树因子,则a1>0, a2>0),再设二元Weibull分布函数对应的概率密度函数为q(x1, x2), 则生存函数可表示为:
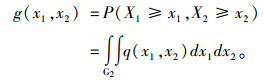
|
(2) |
当x1→a1,

|
(3) |
当x2→a2,
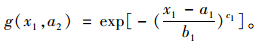
|
(4) |
这样由二元Weibull生存函数结合图 1就有:
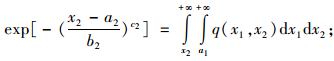
|
(5) |
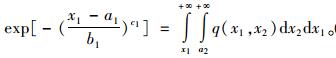
|
(6) |
二元Weibull分布函数为:

|
(7) |
而P(a1≤X1 < +∞, a2≤X2 < +∞)=1, 故二元Weibull分布函数为:
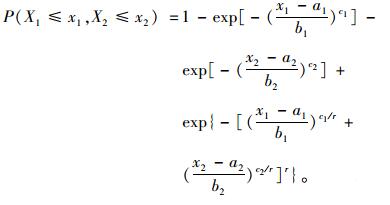
|
(8) |
在(8) 式中令x1→∝, 就得到P(X2≤x2)=1-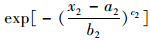
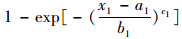
浙江省于1979年建立了森林资源连续清查体系,以5年为一个复查周期。共设置固定样地4 250个,样点格网为4 km×6 km,样地形状为正方形,边长28.28 m,面积800 m2。本研究利用2004年的森林连续清查数据,选择基本为毛竹纯林的样地245个。每个样地毛竹株数18~461不等,样地内毛竹胸径为5~15 cm。龄级划分方法为:当年生竹记为1度竹,2~3年生竹记为2度竹,依此类推。245个样地的数据统计值见文献(刘思斌等,2010)。
2.2 模型参数与评价根据刘思斌等(2010)的累计株数值通过SPSS软件对毛竹各径阶、各龄级实测概率累积值进行估计,非线性最小二乘拟合二元Weibull分布参数a1,b1,c1,a2,b2,c2和r的值分别为1.708, 6.573,3.248, 0.124, 1.427, 1.358和0.989。此时离差平方和为2.728 4×10-4, R2=0.991 0, 这说明模型的估算精度非常高。图 2更直观地描述了二元Weibull分布的测量精度。

|
图 2 省域尺度毛竹胸径、年龄二元Weibull分布图及实测概率散点图 Figure 2 Bivariate Weibull probability distributions and scatter plot of observed probability for diameter-age of moso bamboo on a provincial scale |
从图 2也可以看出:二元Weibull分布函数对浙江省域尺度毛竹胸径、年龄联合分布信息的测量精度非常高。
2.3 各径阶、各龄级毛竹概率由于二元Weibull分布函数估算结果是各径阶、各龄级估计概率的累积值,因此要做相应的减法运算才能得到各径阶、各龄级估计概率的密度值,经相应减法运算得到的毛竹各径阶、各龄级实测概率密度值与对应的估计值(表 1)。
|
|

根据2004年的统计结果,浙江省毛竹总株数为14.566亿株,这样全省毛竹各径阶、各龄级的估算株数见表 2。
|
|
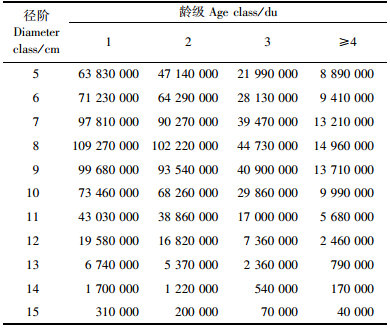
由于表 2所有径阶、龄级毛竹株数的和不等于14.566亿株,所以需要进行修正,其修正方法是表 3的每一数值乘以1.021 054,修正后浙江省各径阶、各龄级毛竹的株数见表 3。
|
|

单株毛竹生物量采用公式(9) 计算,该模型是根据常用生物量模型与毛竹的生长规律,结合浙江省毛竹生物量调查数据建立的, 模型的相关系数R2=0.937, 在0.05置信水平下的预估精度为96.43%, 总系统误差为-0.021%, 符合生物量估算精度要求(周国模, 2006)。
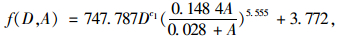
|
(9) |
式中:f(D, A)为毛竹单株生物量(kg),A为单株毛竹年龄(度),D为单株毛竹胸径(cm)。
根据(9) 式可得毛竹单株生物量,结果见表 4。
|
|
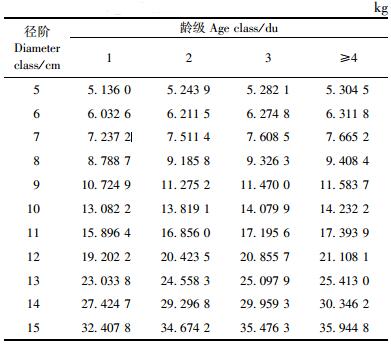
对某区域毛竹生物量进行估算时,抛开中间的换算因子,这样某区域的毛竹总生物量等于该区域所有毛竹单株生物量之和,现用公式(10) 表示如下:

|
(10) |
式中:f(Di, Aj)为第i径阶、第j龄级的毛竹生物量,MT为某区域的总生物量,m为毛竹最大径级值(毛竹的最小径阶值为5),n为毛竹最大龄级值,kij为第i径级、第j龄级的毛竹株数。从(10) 式可以看出:无论是林分尺度还是省域尺度,公式(10) 均适用,因此只要知道任意尺度的毛竹总株数和单株毛竹生物量,就可以准确地推算该尺度的毛竹总生物量。
根据表 3与表 4数据及公式(10),得浙江省各径阶、各龄级的毛竹总生物量(表 5),进而计算出浙江省毛竹总生物量为1.471 6×1010 kg。
|
|
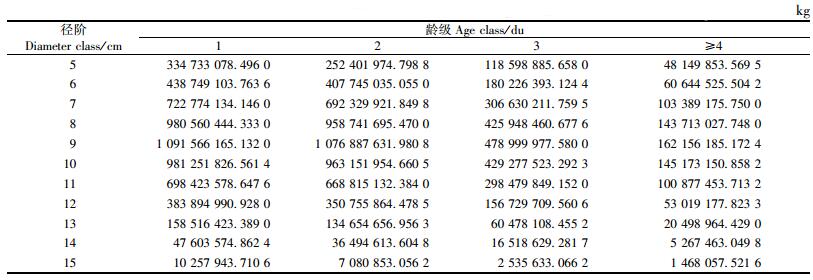
区域尺度生物量的估算一直是研究热点,但目前有关森林生态系统生物量估算的研究多集中在国家尺度或局部典型的森林生态系统上(王效科等, 2001; Isaev et al., 1995; Murillo, 1997; 周玉荣等, 2000; 方晰等, 2002; 周国模等, 2004), 而对中尺度毛竹林生态系统生物量估算的研究相对薄弱。本研究对具有概率模型性质的二元Weibull分布函数做了推导,应用测树因子二元概率分布模型与单株生物量模型,提出了直接精确估算区域生物量的新方法,并用该方法估算出2004年浙江省毛竹总生物量为1.471 6×1010kg,从而实现了首次定量描述浙江省毛竹林的生态功能。
本研究提出的估算区域尺度生物量的方法,弥补了常用生物量尺度转换方法在推算区域生物量时精度不高或不能实现的缺陷,使单株、林分和区域3个不同尺度生物量间的相互转换成为可能,为生物量新尺度转换方法的提出奠定了基础。
本研究提出的方法进还可精确估算区域尺度其他树种的总生物量。应用2个以上测树因子的联合概率分布函数与相应的单株生物量模型,可使区域尺度总生物量的估算结果更精确、更符合实际,但到目前为止有关该方面的报道很少,有待进一步深入研究。
方精云, 刘国华, 徐嵩龄. 1996. 我国森林植被的生物量和净生产量[J]. 生态学报, 16(5): 497-508. |
方晰, 田大伦, 项文化. 2002. 速生阶段杉木人工林碳素密度、贮量和分布[J]. 林业科学, 38(3): 14-19. DOI:10.11707/j.1001-7488.20020303 |
葛宏立, 周国模, 刘恩斌, 等. 2008. 浙江省毛竹直径与年龄的二元Weibull分布模型[J]. 林业科学, 44(12): 15-20. DOI:10.3321/j.issn:1001-7488.2008.12.003 |
刘思斌, 周国模, 施拥军, 等. 2010. 侧树因子二元概率分布:以毛竹为例[J]. 林业科学, 46(10): 29-36. DOI:10.11707/j.1001-7488.20101005 |
史道济, 唐爱丽, 汪玲. 2003. 二元威布尔分布形状参数相等的检验[J]. 天津大学学报, 36(1): 68-71. |
王效科, 冯宗炜, 欧阳志云. 2001. 中国森林生态系统的植物碳储量和碳密度研究[J]. 应用生态学报, 12(1): 13-16. |
赵敏, 周广胜. 2004. 基于森林资源清查资料的生物量估算模式及其发展趋势[J]. 应用生态学报, 15(8): 1468-1472. |
周国模. 2006. 毛竹林生态系统中碳储量、固定及其分配与分布的研究[J]. 浙江大学博士学位论文. |
周国模, 姜培坤. 2004. 毛竹林的碳密度和碳储量及其空间分布[J]. 林业科学, 40(6): 20-24. DOI:10.11707/j.1001-7488.20040604 |
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳贮量和碳平衡[J]. 植物生态学报, 24(5): 518-522. |
Brown S. 2002. Measuring carbon in forests: current status and future challenges[J]. Environmental Pollution, 116(3): 363-372. DOI:10.1016/S0269-7491(01)00212-3 |
Brown S, Lugo A E. 1984. Biomass of tropical forests: a new estimate based on forest volumes[J]. Science, 223(4642): 1290-1293. DOI:10.1126/science.223.4642.1290 |
Dixon R K, Solomon A M, Brown S, et al. 1994. Carbon pools and flux of global forest ecosystems[J]. Science, 263(5144): 185-190. DOI:10.1126/science.263.5144.185 |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 292(5525): 2320-2322. DOI:10.1126/science.1058629 |
Isaev A, Korovin G, Zamolod D. 1995. Carbon stock and deposition in phytomass of the Russian forests[J]. Water Air Soil Poll, 82(1/2): 247-256. |
Johnson W C, Sharpe D M. 1983. The ratio of total to merchantable forest biomass and its application to the global carbon budget[J]. Can J For Res, 13(3): 372-383. DOI:10.1139/x83-056 |
Murillo J C R. 1997. Temporal variations in the carbon budget of forest ecosystems in Spain[J]. Ecological Applications, 7(2): 461-469. DOI:10.1890/1051-0761(1997)007[0461:TVITCB]2.0.CO;2 |
Shi D J. 1995. Moment estimation for multivariate extremes value distribution[J]. Appl Math-JCU, 10(1): 61-68. DOI:10.1007/BF02663895 |
Tawn J A. 1988. Bivariate extreme value theory:models and estimation[J]. Biometrika, 75(3): 397-415. DOI:10.1093/biomet/75.3.397 |
Zhou G S, Wang Y H, Jiang Y L. 2002. Estimating biomass and net primary production from forest inventory data: a case study of China's Larix forests[J]. Forest Ecology and Management, 169(1/2): 149-157. |
Zhang X Q, Xu D Y. 2002. Calculation forest biomass change in China[J]. Science, 296(5572): 1359. DOI:10.1126/science.296.5572.1359a |