 2010, Vol. 46
2010, Vol. 46文章信息
- 韩广轩, 王光美, 毛培利, 张志东, 于君宝, 许景伟
- Han Guangxuan, Wang Guangmei, Mao Peili, Zhang Zhidong, Yu Junbao, Xu Jingwei
- 山东半岛北部黑松海防林幼龄植株更新动态及其影响因素
- Regeneration Dynamics of Young Pinus thunbergii and Its Influencing Factors in the Coastal Protection Forests in Northern Shandong Peninsula
- 林业科学, 2010, 46(12): 158-164.
- Scientia Silvae Sinicae, 2010, 46(12): 158-164.
-
文章历史
- 收稿日期:2009-04-13
- 修回日期:2010-10-29
-
作者相关文章



2. 山东省林业科学研究院 济南 250014
2. Shandong Provincial Academy of Forestry Jinan 250014
海防林是人工林生态网络系统的重要组成部分,也是海岸带地区典型的生态系统类型。印度洋海啸发生后,中国更加重视沿海防护林体系建设,制定了《全国沿海防护林体系建设工程规划(2006— 2015年)》,规划土地总面积为4 471万hm2,占国土总面积的4.7%,总投资为99.84亿元,其中包括开展沿海防护林科学研究等科技支撑体系建设(国家林业局,2008)。可见,目前人们对海防林的重要性和科学研究的必要性有了深刻认识。
由于海护林处在海陆相互作用活跃的过渡地带,风害、盐害、雾害、水淹等自然灾害频繁发生,造林比较困难,因此,如何使现有防护林的防护效益不间断,仍是目前海防林经营的一个难点(朱教君等,2002)。森林天然更新是森林资源再生产的一个自然生物学过程,对森林生态系统生产力、未来森林群落结构及其生物学多样性有着重要影响,因而是实施森林可持续经营过程中最重要的环节(宋新章,2007)。由于天然更新充分依靠自然的力量来实现森林生态环境的自然恢复,并且能够培育出合乎自然规律的高生物多样性和高生态质量的森林(刘足根等,2007)。因此,现有海防林成功地实现天然更新将是最理想的。
森林树种幼龄植株更新是森林动态变化的关键阶段,在森林树种生活史中,幼龄植株阶段是对环境条件反应最敏感的时期(韩有志等,2002; Du et al., 2007)。树木幼龄植株更新受生物因素和非生物因素综合作用的影响,例如更新树种的遗传学、生理学、生态学特性及其与周围树种之间的关系(如植物种间的竞争、化感作用)、幼龄植株所处的生境条件以及自然和人为干扰的类型、尺度、强度、频率等都会影响林木幼龄植株更新(李小双等,2007; 朱教君等,2008)。因此,在林木天然更新过程中,幼龄植株的定居和生长发育是林木种群维持和实现更新的一个重要阶段(Guariguata,2000),对林木更新具有重要的筛选作用(Rey et al., 2000)。由于树木幼龄植株能决定树木种群在群落演替系列中的位置以及成年树木种群在群落中的地位(陶建平等,1997),因此,对树木幼龄植株的生长规律、种群动态及其影响因素进行研究具有重要的理论和实际意义。
黑松(Pinus thunbergii)具有喜海洋气候,适应性强,抗海风,耐瘠薄,在中性或微碱性砂滩、海岸能良好生长等特点(朱教君等,2002)。20世纪50年代在我国山东半岛和辽东半岛大面积种植黑松,目前黑松已成为山东半岛海岸基干林带的主要造林树种,面积约为7万hm2,占山东省海防林面积的70%以上(许景伟等,2005)。黑松经过半个多世纪引种,有些地域现已自然更新成林,但是在成林过程中,幼龄植株生长动态和种群数量是怎样变化的?影响黑松幼龄植株更新的主要因素是什么?目前这方面的研究尚不多见。本文以我国山东半岛北部黑松海防林为研究对象,分析黑松幼龄植株的数量特征及生长动态,探讨林分条件(成年乔木、灌木层和草本层)对黑松幼龄植株更新的影响,不仅有助于阐明黑松海防林的天然更新过程与机制,而且为黑松海防林的保育和可持续经营提供科学依据。
1 研究区概况研究区位于中国科学院烟台海岸带可持续发展研究所临海台站(121°41′57″ E,37°27′15″ N),属暖温带东亚季风型大陆性气候,年平均温度11.5℃,降水量760 mm,无霜期200天左右,年平均相对湿度为71%,年平均日照时数为2 642.7 h,日照率为60%。研究区海拔1 ~ 2 m,地势平坦,坡度小于5°,土壤为滨海砂地土壤,机械组成以砂为主,有机质含量很低,一般不超过1%。黑松林栽植于20世纪50年代,大多为纯林,黑松种群在林分中占居主林层,为优势树种; 幼苗和幼树占相当大的比例,Ⅰ(H < 0.33 m)和Ⅱ(0.33 m≤ H < 1.5 m,DBH < 2.5 cm)径级的黑松分别占35.4%和16.0%,老龄级株数缺乏,DBH > 7.5 cm径级的黑松不到种群的0.01%。该样地林分状况详见韩广轩等(2008)。黑松林下植被比较简单,灌木主要由紫穗槐(Amorpha fruticosa)、单叶蔓荆(Vitex trifolia var.simplicifolia)、胡枝子(Lespedeza bicolor)、野蔷薇(Rosa multiflora)等树种组成; 草本层主要为羊胡子草(Carex rigescens)、虫实(Corispermum sp.)、刺沙蓬(Salsola ruthenica)、白茅(Imperata cylindrica var.major)、马齿苋(Portulaca oleracea)、肾叶打碗花(Calystegia soldanella)等。
2 研究方法 2.1 调查方法在垂直于海岸线方向,每隔200 m设置1条样线,共3条样线。沿每条样线每隔50 m,设置10 m × 10 m的样方1个,每条样线14个样方,共设置42个样方。在每个样方中对乔木进行每木检尺,记录其高度、胸径(DBH)、冠幅等特征,其中冠幅利用皮尺量测,通过量取树冠投影的长度以及与长度相垂直投影的树冠宽度获得。高度低于1.5 m的黑松幼龄植株记录其高度、基径、数量及其与最近立木的距离,并采用查数轮生枝法确定其年龄(苗木年龄小,轮枝清楚,较为准确)。同时记录灌木的高度、基径、丛径等。在每个10 m × 10 m乔木样方内选择4个顶点及对角线中心点调查5个1 m × 1 m的草本样方,调查草本层的物种名、冠径、丛径、盖度等。
2.2 种群更新状况测定采用查数轮生枝法确定幼龄植株年龄,统计每一样方各龄级的株数,将所有样方各龄级的株数合并,组成黑松幼树种群年龄结构的基本数据; 以0.1 m为一高度级,统计各高度级内幼龄植株的株数以确定高度结构; 以0.3 m为一距离级,统计各距离级内黑松幼龄植株的株数以确定幼龄植株与最近立木的距离。分别绘制黑松幼龄植株种群龄级、高度级、距离级与个体密度分布图,以分析其种群结构变化。
2.3 数据处理运用非线性回归方法分析黑松幼龄植株平均树高、平均基径和密度与年龄的关系; 运用线性回归方法分析黑松幼龄植株种群平均高度与平均基径的关系; 运用非线性回归方法分析黑松幼龄植株距最近立木的空间分布模式; 运用相关分析方法分析幼龄植株的密度、高度、基径和苗龄与林分条件(成年乔木、灌木层和草本层)之间的关系。采用SPSS 12.0统计分析软件进行数据统计分析,绘图在Sigmaplot 9.0软件中完成。
3 结果与分析 3.1 不同样方黑松幼龄植株的密度和数量研究区不同样方黑松幼龄植株的一些基本数量特征参见表 1。通过对42块样方的天然黑松更新调查可知,黑松以种子萌发更新为主。各样方黑松更新密度在50 ~ 24 300株·hm-2之间波动,平均更新密度为4 024株·hm-2,接近一般更新造林的标准4 444株· hm-2 (孙景波等,2009)。超过38%的样方更新株数在2 000 ~ 5 000株·hm-2,30%的样方黑松更新株数 > 4 000株·hm-2,超过30%的样方黑松更新株数在1 000株·hm-2以下。各个样方中的黑松种群所处的生境及母树数量、结实状况不同,致使各样方在种子数量、萌发及幼苗建立等方面也不相同,造成各样方黑松幼龄植株种群密度上的较大差异,变异系数为127.1%。
|
|

黑松幼龄植株高生长和基径生长具有相同的趋势,幼龄植株树高和基径与年龄均呈指数函数关系(图 1)。幼龄植株高生长速度为0.01 ~ 0.19 m·a-1,幼龄植株在4年生以前生长缓慢,4年生幼龄植株平均树高仅为0.15 m; 从5年生开始,幼龄植株高生长加快,11年生幼龄植株生长速度达到0.19 m·a-1,在12年生时,幼龄植株平均树高达1.05 m。幼龄植株基径生长速度为0.03 ~ 0.33 cm·a-1,在4年生以前基径生长缓慢,生长速率为0.04 cm·a-1; 从5年生开始,幼龄植株基径生长加快,在0.11 ~ 0.33 cm·a-1,在12年生时,幼龄植株平均基径达2.01 cm。另外,在黑松更新阶段,幼龄植株的树高与基径之间呈直线回归函数相关(图 2),即y (树高) = 0.503 6x (基径) + 0.001 9(R2 = 0.99),反映幼龄植株还处在高生长阶段(何兴元等,2003)。

|
图 1 黑松幼龄植株树高和基径随年龄变化的生长动态 Figure 1 Growth dynamics of individual height and basal diameter of young P. thunbergii trees with age change |

|
图 2 黑松幼龄植株高度与基径的关系 Figure 2 Relationship between height and basal diameter of young P. thunbergii trees |
以年龄为横坐标,各龄级幼龄植株的密度为纵坐标,绘制黑松幼龄植株的林龄结构图(图 3)。由图 3可见,黑松幼龄植株的林龄结构呈明显的偏态分布。黑松幼龄植株中的幼龄个体较少,1 ~ 2年生植株占总数的9.57%。而在3 ~ 7年这个年龄段存活株数达到最高,占总数的76.39%;从8年之后,存活株数明显减少。黑松3 ~ 7年植株个体在数量上表现出优势地位,说明该黑松幼龄植株是处于一种相对稳定的状态,而随着年龄增长幼龄植株数量进一步下降。

|
图 3 黑松幼龄植株的年龄结构 Figure 3 Age structure of young P. thunbergii trees |
由于1 ~ 2生幼龄植株的种群命运很不确定,受环境的影响很大,从发生到死亡都有不确定性(杨玲,2007)。因此,只分析3 ~ 12年生黑松幼龄植株密度与年龄的相关关系。回归分析表明,3 ~ 12年生黑松幼龄植株密度与年龄之间呈指数相关关系:
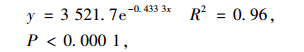
|
(1) |
式中,y为密度(株·hm-2),x为年龄(a)。
在年龄与密度关系中(图 3),黑松幼龄植株密度从3龄的989株·hm-2下降到12龄的35株· hm-2,幼龄植株密度随着年龄的增加而呈指数级下降,这可能是由于随着个体增长,对环境条件的需求增大,造成较大的种内竞争,使得黑松幼龄植株在发育过程中同龄个体间产生明显的自疏现象。
3.4 幼龄植株高度结构以0.1 m为一个高度级,对黑松幼龄植株高度结构进行统计,结果见图 4。黑松幼龄植株高度结构表现为金字塔型,个体数量随着高度级的增加而减少(图 4)。0.1 ~ 0.2 m的黑松个体数量最多,占总株数的44.9%,其次为0 ~ 0.1 m的黑松个体,占总株数的18.7%。黑松幼龄植株更新株数主要集中在0 ~ 0.4 m,其间黑松个体比例高达82.2%。

|
图 4 黑松幼龄植株的高度结构 Figure 4 Height structure of young P. thunbergii trees |
黑松幼龄植株距最近立木的距离表现出两端低、中间高的“山峰型”分布曲线(图 5),回归分析表明,其分布格局呈高斯分布:

|
(2) |

|
图 5 黑松幼龄植株与最近立木的距离 Figure 5 Distance of young P. thunbergii trees away from the nearest standing tree |
式中,y为密度(株·hm-2),x为与最近立木的距离(m)。
黑松幼龄植株主要集中分布在距最近立木0.3 ~ 0.9 m的范围内,这一范围内的更新总数占全部更新株数的74.7%,离立木越近或越远幼龄植株均逐渐减少。距最近立木≤0.3 m的范围内,黑松幼龄植株占全部更新株数的7.80%;幼龄植株离立木的最远距离为3.3 m,但距最近立木 > 2.1 m的范围内,黑松幼龄植株仅占全部更新株数的0.39% (图 5)。
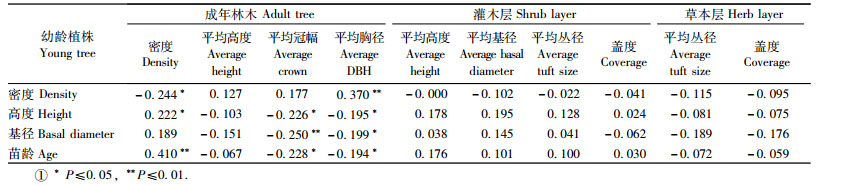
3.6 幼龄植株更新的影响因素由于黑松海防林土壤以滨海砂地土壤为主,土层较薄,土壤养分含量均较低,加之该研究区地势平坦,无坡度和坡向等地形因子的影响,所以,本文主要观测林分条件(成年乔木、灌木层和草本层)对黑松幼龄植株更新的影响。相关分析表明,黑松幼龄植株更新主要受成年乔木(H > 1.5 m)的影响,而与灌木层和草本层的生长状况无关。由表 1可以看出,幼龄植株密度主要受乔木密度和乔木平均胸径的影响; 幼龄植株高度与乔木密度、乔木平均冠幅和胸径之间的相关系数均达到显著水平(P≤0.05);幼龄植株基径与乔木平均冠幅之间呈极显著负相关(P≤0.01),与乔木平均胸径之间呈显著负相关(P≤0.05);幼龄植株年龄与乔木密度之间呈极显著正相关(P≤0.01),与乔木平均冠幅和胸径均呈显著负相关(P ≤ 0.05)。但是,乔木平均高度对黑松幼龄植株更新的影响不显著。灌木(平均高度、基径、丛径和盖度)和草本(平均丛径和盖度)的盖度与黑松更新状况之间并没有明显的相关关系(P > 0.05),说明灌木和草本生长状况不影响黑松幼龄植株的更新。
|
|
研究种群林龄结构不仅可以了解现在的种群状态,也可以分析过去种群的结构及受干扰状态,预测未来的种群动态,并且有助于揭示种子散布、萌发及幼苗建立等特征(谢宗强等,1999)。从林龄结构来看,黑松幼龄植株的林龄结构呈明显的偏态分布,1 ~ 2年生幼龄植株为“倒金字塔”型结构,3年生种群数量最大,4年生以后,幼龄植株种群数量随年龄的增加而减小。以种子繁殖为主的植物种群在缺少幼龄植株时,一般判断为衰退型种群(Manuel,2000)。但实际上,这些1 ~ 2生幼龄植株的命运很不确定,受环境的影响很大,从发生到死亡都有不确定性(杨玲,2007)。从8年之后,黑松的存活株数明显减少。相对来说,3 ~ 7年的幼龄植株个体对确定幼龄植株种群未来命运是至关重要的,因为这个阶段的个体处于种群的建立阶段(杨玲,2007)。在3 ~ 7年这个年龄段,黑松幼龄植株存活株数达到最高,占总数的76.39%,在数量上表现出优势地位,有理由判断该黑松幼龄植株种群是处于一种相对稳定的状态。
4.2 黑松幼龄植株的空间分布从空间分布上看,幼龄植株主要集中分布在距最近立木0.3 ~ 0.9 m的范围内,这一范围内的更新总数占全部更新株数的74.7%,离立木越近或越远幼龄植株均逐渐减少(图 5)。黑松幼龄植株距最近立木的距离表现出两端低、中间高的“山峰型”分布曲线。黑松幼龄植株的这种分布规律与前人的研究结果相似。例如,与母树的距离极大地影响鼎湖山林窗内的幼树密度(彭闪江等,2003); 天然红树林中的海桑(Sonneratia caseolaris)幼龄植株扩散受到与母树距离的影响显著,离母树越远,幼龄植株的密度越小(曾雪琴等,2008)。
幼龄植株和母树以及距离之间的制约关系非常复杂,造成制约的原因可能与母树周围的种子密度、幼苗竞争、微环境的空间差异等因素有关。一方面,距离母树远近,种子密度存在差异。例如,邹莉等(2007)研究发现,从种子雨的扩散强度来看,神农架巴山冷杉(Abies fargesii)种子雨雨量随着与中心母树距离的增加而迅速减少,呈现出近似正态分布。另一方面,距离母树不同,种子被取食者取食或被病原体侵染的概率也不同。杜彦君等(2007)对鼎湖山针阔混交林的研究表明被病原体侵染的锥栗(Castanopsis chinensis)种子的命运受距离制约。另外,与最近立木的距离不同,相应的微环境也发生变化,林内的光照、地面植被和土壤性质等生境条件可能会表现出明显的斑块性,这种小尺度上生境条件的空间异质性特征可能影响树苗的定居,影响到幼龄植株的空间分布。
4.3 林分条件对黑松幼龄植株更新的影响森林天然更新是一个复杂的生态学过程,无论是有性更新还是无性更新,都受到环境梯度的刻划(梁建萍等,2002; Zhu et al., 2003)。例如,人工林天然更新就受林分密度、林缘效应、种源、立地、林地形状与面积大小等多种因素的干扰(孙景波等,2009)。本文主要观测林分条件(成年乔木、灌木层和草本层)对黑松幼龄植株更新的影响。结果表明,黑松幼龄植株更新主要受成年乔木密度、平均冠幅和胸径的影响,而与乔木平均高度无关; 同时,灌木层和草本层的生长状况对黑松幼龄植株更新的影响也不显著。由于乔木密度、平均胸径和冠幅是反映林冠层疏密程度的主要因素,因此,林冠层的疏密程度可能是影响黑松幼龄植株更新的主要因素。黑松幼龄植株更新密度与成年乔木密度呈显著负相关,说明随着林分密度或郁闭度的增加,黑松幼龄植株的密度逐渐减少。黑松为不耐荫树种,林冠层的疏密程度直接影响光照条件和营养面积,从而影响种子的产量和质量、种子萌发和幼苗定居过程(徐鹤忠等,2006)。例如,郁闭的云冷杉林在林冠下更新的数量、质量往往不及较稀疏的林分(韩景军等,2000); 郁闭度较大的樟子松(Pinus sylvestris var. mongolica)林内未发现有更新的幼苗(曾德慧等,2002)。
另外,林下的灌草层也是一种重要的生态学滤筛,它可以改变微生境的条件,如地表光照、温度、土壤湿度等,改变凋落物的分布,还间接影响种子取食者的行为,从而对幼苗的定居产生影响(彭闪江等,2004; 汤景明等,2005)。但是,本文研究结果表明,黑松幼龄植株更新状况与灌木层和草本层的生长状况无关,这一结果与Sharew等(1997)和刘足根等(2007)发现草本和灌木不影响非洲圆柏(Juniperus procera)和长白落叶松(Larix olgensis)幼苗萌发的结果相似。这可能是由于黑松海防林大多为纯林,林下植被比较简单,如灌木种类主要有紫穗槐、单叶蔓荆、胡枝子、野蔷薇等,且数量稀少,平均密度仅为2 184.0株·hm-2,可能对黑松幼龄植株的定居和生长影响不明显。据以上分析,黑松海防林更新管理应该充分考虑对成年乔木对幼龄植株的影响,对浓密的成年乔木实行间伐等抚育措施,保留适当密度,既不减少其防护效能,又能为林下幼龄植株更新创造良好的光照条件,以利于黑松幼龄植株定居和生长,促进黑松海防林天然更新。
杜彦君, 彭闪江, 徐国良, 等. 2007. 鼎湖山针阔混交林锥栗种子距离制约研究[J]. 植物生态学报, 31(6): 998-1006. |
国家林业局. 2008. 全国沿海防护林体系建设工程规划简介. http://www.forestry.gov.cn/distribution/2008/04/22/jhyzj-2008-04-22-1460.html.
|
韩广轩, 王光美, 张志东, 等. 2008. 烟台海岸黑松防护林种群结构及其随离岸距离的变化[J]. 林业科学, 44(10): 8-13. DOI:10.3321/j.issn:1001-7488.2008.10.002 |
韩景军, 肖文发, 罗菊春. 2000. 不同采伐方式对云冷杉林更新与生境的影响. 2000[J]. 林业科学, 36(专1): 90-96. |
韩有志, 王政权. 森林更新与空间异质性[J]. 应用生态学报, 13(5): 615-619. |
何兴元, 陈玮, 徐文铎, 等. 2003. 城市近自然林的群落生态学剖析:以沈阳树木园为例[J]. 生态学杂志, 22(6): 162-168. |
李小双, 彭明春, 党承林. 2007. 植物自然更新研究进展[J]. 生态学杂志, 26(12): 2081-2088. |
梁建萍, 王爱民, 梁胜发. 2002. 干扰与森林更新[J]. 林业科学研究, 15(4): 490-498. |
刘足根, 朱教君, 袁小兰, 等. 2007. 辽东山区长白落叶松天然更新调查[J]. 林业科学, 43(1): 42-49. |
彭闪江, 黄忠良, 周国逸, 等. 2003. 鼎湖山林窗形成特征及其对幼树组成和多样性的影响[J]. 热带亚热带植物学报, 11(3): 229-235. |
彭闪江, 黄忠良, 彭少麟, 等. 2004. 植物天然更新过程中种子和幼苗死亡的影响因素[J]. 广西植物, 24(2): 113-121. |
宋新章. 2007. 长白山区采伐林隙更新及其微生境研究. 中国林业科学研究院博士学位论文. http://d.wanfangdata.com.cn/Thesis/Y1173950
|
孙景波, 佟静秋, 牟长城, 等. 2009. 哈尔滨城市人工林天然更新组成结构与年龄结构[J]. 东北林业大学学报, 37(2): 16-21. |
汤景明, 翟明普. 2005. 影响天然林树种更新因素的研究进展[J]. 福建林学院学报, 25(4): 379-383. |
陶建平, 钟章成. 1997. 不同环境中四川大头茶幼苗的发生及幼苗消亡过程的研究[J]. 西南师范大学学报:自然科学版, 22(3): 249-256. |
谢宗强, 陈伟烈, 路鹏, 等. 1999. 濒危植物银杉的种群统计与年龄结构[J]. 生态学报, 19(4): 523-528. |
徐鹤忠, 董和利, 底国旗, 等. 2006. 大兴安岭采伐迹地主要目的树种的天然更新[J]. 东北林业大学学报, 34(1): 18-21. |
许景伟, 李传荣, 王卫东, 等. 2005. 沿海沙质岸黑松防护林的生物量及生产力[J]. 东北林业大学学报, 33(6): 29-32. |
杨玲. 2007. 天童常绿阔叶林萌枝更新的生态学研究. 华东师范大学硕士学位论文, 28-33. http://cdmd.cnki.com.cn/article/cdmd-10269-2007081111.htm
|
曾德慧, 尤文忠, 范志平, 等. 2002. 樟子松人工固沙林天然更新障碍因子分析[J]. 应用生态学报, 13(3): 257-261. |
曾雪琴, 陈鹭真, 谭凤仪, 等. 2008. 深圳湾引种红树植物海桑的幼苗发生和扩散格局的生态响应[J]. 生物多样性, 16(3): 236-244. |
朱教君, 刘足根, 王贺新. 2008. 辽东山区长白落叶松人工林天然更新障碍分析[J]. 应用生态学报, 19(4): 695-703. |
朱教君, 李凤芹, 松崎健, 等. 2002. 间伐对日本黑松海岸林更新的影响[J]. 应用生态学报, 13(11): 1361-1367. DOI:10.3321/j.issn:1001-9332.2002.11.001 |
邹莉, 谢宗强, 李庆梅, 等. 2007. 神农架巴山冷杉种子雨的时空格局[J]. 生物多样性, 15(5): 500-509. |
Du X J, Guo Q F, Ma K P, et al. 2007. Seed rain, soil seed bank, seed loss and regeneration of Castanopsis fargesii (Fagaceae) in a subtropical evergreen broad-leaved forest[J]. Forest Ecology and Management, 238: 212-219. DOI:10.1016/j.foreco.2006.10.018 |
Guariguata M R. 2000. Seed and seeding ecology of tree species in neotropical secondary forest: management implications[J]. Ecological Applications, 10(1): 145-154. DOI:10.1890/1051-0761(2000)010[0145:SASEOT]2.0.CO;2 |
Manuel C M. 2000. Ecology: Concepts and application[M]. Beijing: Science Press.
|
Rey P J, Alcántara J M. 2000. Recruitment dynamics of a fleshy-fruited plant (Oleaeuropaea) : connecting patterns of seed dispersal to seedling establishment[J]. Journal of Ecology, 88: 622-633. DOI:10.1046/j.1365-2745.2000.00472.x |
Sharew H, Legg C J, Grace J. 1997. Effects of ground preparation and microenvironment on germination and natural regeneration of Juniperus procera and Afrocarpus gracilior in Ethiopia[J]. Forest Ecology Management, 93: 215-225. DOI:10.1016/S0378-1127(96)03962-X |
Zhu J J, Matauzaki T, Li F Q, et al. 2003. Effects of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest[J]. Forest Ecology and Management, 182: 339-354. DOI:10.1016/S0378-1127(03)00094-X |