 2010, Vol. 46
2010, Vol. 46文章信息
- 邓磊, 张文辉
- Deng Lei, Zhang Wenhui
- 黄土沟壑区刺槐人工林的天然发育规律
- Natural Development Pattern of Robinia pseudoacacia Plantations in Loess Hilly Region
- 林业科学, 2010, 46(12): 15-22.
- Scientia Silvae Sinicae, 2010, 46(12): 15-22.
- DOI: 10.11707/j.1001-7488.20101203
-
文章历史
- 收稿日期:2009-07-28
- 修回日期:2010-09-17
-
作者相关文章



2. 西北农林科技大学西部环境与生态教育部重点实验室 杨凌 712100
2. Key Laboratory of Environment and Ecology in West China of Education Ministry, Northwest A & F University Yangling 712100
人工植被天然化发育是指人工植被建成后,建群种种群能实现自我更新,随着时间的推移,乡土物种入侵,灌草层形成,群落功能持续发育的生态学过程。一般情况下,只有完成造林或播种过程,建群种达到一定成活率,当地的乡土植物、动物和微生物就会很快进入,参与群落的形成与演化。黄土高原地区人工林建设应该以生态效益为第一目标,栽植的建群种能够通过有性或者无性繁殖,实现自我更新,形成可以持续利用的人工生态系统,避免周期性衰退,就能实现永续利用(张文辉等,2009)。
刺槐(Robinia pseudoacacia)是黄土高原地区人工林面积最大的树种。刺槐人工群落在黄土高原地区年降水量>550 mm地区,通过根系萌苗、伐桩萌苗可以实现群种群恢复和群落更新。但在降水量<550 m丘陵沟壑区,有些生境自我更新困难。近年来,很多学者对刺槐在黄土高原丘陵沟壑区是否适合营造人工林提出质疑(韩蕊莲等,2003)。由于缺水,刺槐造林后出现永久性“土壤干层”(杨文治,2001),林木个体发育迟缓,形成了“小老树”。特别是在阳坡生境问题更为严重,感染病虫,自然枯死,林木几乎没有经济价值,并且恶化土壤水分条件。这些似乎表明丘陵沟壑区不适合刺槐人工林栽培。但是,据观测,在丘陵沟壑区阴坡下部生境,刺槐人工林通过根系萌苗、伐桩萌苗等无性繁殖方式自我更新,具有天然化培育的潜力。以前对黄土高原刺槐人工群落研究主要集中在群落特征、林地土壤养分水分、个体生长发育等方面(曹扬等,2006),而对丘陵沟壑区阴坡阳坡不同发育阶段刺槐人工林群落特征、种群年龄结构等方面对比研究不多。针对丘陵沟壑区阴坡阳坡生境,以种群能否更新,群落能否持续发育研究未见报道,而这正是刺槐人工林能否天然化培育的关键。如果刺槐人工林建群种种群无性繁殖的幼苗数量足够多,能实现自我更新,林内物种多样性和水分养分足够丰富,说明其符合天然化培育条件,否则,可以考虑不再在此地营造刺槐人工林。
本文选择黄土高原丘陵沟壑区20世纪60年代以来造林的刺槐人工林,通过样地调查,分析建群种径级结构、高度级结构、林下物种多样性、土壤养分水分等,比较阴坡与阳坡在不同发育阶段刺槐人工种群发育及群落特征演化,目的是阐明刺槐林在不同生境条件下种群更新潜力、群落特征和生态功能发育过程,为黄土丘陵沟壑区刺槐人工林天然化发育的生境选择及经营管理提供依据。
1 研究地区与研究方法 1.1 研究地区自然概况研究区域选择在中国科学院水土保持研究所水土保持综合治理试验区陕西省安塞县纸坊沟、延安宝塔区柳林镇,属黄土高原腹地,地理位置36°51′- 37°02′ N,109° 19′-110° 07′ E,海拔1 010 ~ 1 431 m。属暖温带半干旱气候区,年平均降水量为510 ~ 530 mm,7-9月占全年降水的55% ~ 58%,年均气温8.8 ℃,≥10 ℃年积温2 800 ~ 3 500 ℃。本区为森林区向森林草原的过渡地段。侧柏(Platycladus orientalis)、辽东栎(Quercus liaotungensis)林等斑块状分布于阴坡生境。天然灌木草本群落主要分布于人为干扰较少的沟坡、峁顶或远离居民点石质山地。天然灌丛群落地主要种类有达乌里胡枝子(Lespedeza dahurica)、树锦鸡儿(Caragana arborescens)、扁核木(Prinsepia uniflora)、丁香(Syzygium aromaticum)、悬钩子(Rubus corchorifolius)。组成草地群落的主要种类有铁杆蒿(Artemisia gemlinii)、茭蒿(Artemisia giraldii)、长芒草(Stipa bungeana)、铁线莲(Clematis fruticosa)、艾蒿(Artemisia argyi)等(韩蕊莲等,2003; 侯庆春等,2000;王义凤,1991; 张仰渠,1989)。人工林主要树种有刺槐、油松(Pinus tabulaeformis)。本地区土壤主要为黄绵土。
1.2 研究方法 1.2.1 样地调查在阴坡、阳坡选择1968年(40年)、1984年(25年)和1999年(10年) 3个时间段营造的刺槐人工林。造林措施及后期管理历史清楚。当地刺槐林营造一般采用台阶地基础上穴状整地; 当地培育的2年生实生苗,高度平均35 cm,根系完好; 春季雨后造林,造林株行距1.5 m × 1.5 m。为了保证成活率,在造林后第2年对未成活幼苗进行补种,从而减少阳坡和阴坡造林成活率差异; 造林后禁止放牧、砍伐,避免人为干扰。
在每个生境和时间段选择生长发育较好地段,设置样地3块,共完成样地18块。样地调查时间在刺槐林生长发育旺盛的7-8月进行。样地面积为20 m × 20 m,每个样地分别沿对角线设置3个5 m × 5 m的灌木样方和3个1 m × 1 m的草本样方。调查内容包括: 1)群落学特征:造林时间、造林密度、群落总郁闭度、平均高度; 2)刺槐种群调查:个体坐标定位(以样地一条边为X轴,垂直边为Y轴),测定高度、胸径、冠幅、枯梢率,特别注意幼苗幼树个体起源(根系萌苗、伐桩萌苗)及其生长状况; 3)灌木草本样方调查:分物种测定个体数量、高度、盖度、频度、基径和冠幅; 4)人为干扰:包括砍伐、放牧、开垦等,记录其伐桩数、枝条受损数占样地总数的比例,以及地面扰动程度,强度最大赋值1,强度最小赋值0; 5)土壤养分、水分取样:在样地调查结束后,选择此前10天无雨天气,集中在2天内完成土壤养分测定取样,4天内完成土壤水分取样。取样期间无雨。
1.2.2 刺槐人工林种群统计径级(size class,SC)的划分方法为:以胸径(DBH)2 cm为1个径级,共划分10个径级,分别用Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ,Ⅵ,Ⅶ,Ⅷ,Ⅸ,Ⅹ表示。然后按样地统计每一径级的个体数量(幼苗以基径为准),将同一生境(坡向)样地数据合并,绘制径级结构图。高度级(height class,HC)的划分方法为:以树高(tree height,TH)2 m为1个高度级,共划分10个高度级,也分别用Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ,Ⅵ,Ⅶ,Ⅷ,Ⅸ,Ⅹ表示。然后按样地统计每一高度级的个体数量,将同一生境样地数据合并,绘制高度级结构图(张婷等,2007)。
1.2.3 群落物种多样性测度的方法以样地为单位,分别统计造林10,25和40年刺槐人工林个体特征、林分特征和繁殖特性,分阳坡、阴坡,将相似生境样地数据合并。根据样地调查所得不同物种相对盖度、相对多度和相对频度计算林下物种的重要值,再统计出同一种群中各物种的平均重要值; 分层(灌、草)计算各物种的α多样性指数,如物种丰富度指数(S)、物种多样性指数Simpson指数(D)、Shannon-Wiener指数(H′)、均匀度指数Pielou指数(Jsw)和Alatalo指数(Ea),具体计算方法见文献(蒋志刚等,1997;马克平等,1994)。
1.2.4 土壤水分和养分的测定土壤水分、养分的取样是沿样地内对角线各取3个0 ~ 20 cm土壤混合样品。深层土壤水分测定是在不同发育阶段不同生境的样地内,土钻取土,分别从地表每隔20 cm取1个土样,深度取到200 cm处; 在200 cm以下,每隔50 cm取1个土样,取到400 cm处。采集的土样带回室内测定分析。其中土壤水分含量的测定采用烘箱(85 ℃)法; 土壤pH值应用ZD-2型电位滴定计测定; 有机质用重铬酸钾-H2SO4容量法进行测定; 有效磷应用NaHCO3浸提-钼蓝比色法; 速效氮应用碱解扩散法(阮成江等,2002; 卢金伟等,2002); 速效钾采用火焰光度法(李立平等,2003)。测定数据按不同生境进行样地合并,取平均值。
2 结果与分析 2.1 不同发育阶段下阴坡与阳坡刺槐人工林种群动态 2.1.1 刺槐人工林种群径级结构人工林建群种种群径级结构可以反映种群更新潜力以及同龄个体生长分化和其对生境的适合程度(张婷等,2007)。图 1表明:造林后10,25,40年3个年龄段,阴坡刺槐种群都有一定数量萌生幼龄个体,而阳坡种群越向后期,萌生幼龄个体数量越少。人工林的种群个体是同龄的。在10年生阶段,2种生境幼龄个体数量差异不大,Ⅰ,Ⅱ径级个体数量可能是植苗造林个体生长分化,而不是萌生幼苗。经过25年和40年生的林分发育,Ⅰ,Ⅱ径级幼苗属于萌生幼苗,大于Ⅳ的径级的个体属于植苗造林的个体。在阳坡生境,幼苗数量很少,0.5 ~ 1株·(10 m)-2; Ⅵ径级数量最多。在阴坡生境,幼苗数量较多,3 ~ 3.25株·(10 m)-2; Ⅶ径级数量最多。这说明阴坡生境有利于个体生长发育,不仅植苗造林的个体直径大,而且种群具有一定数量的萌生幼苗; 而阳坡生境刺槐人工种群更新幼苗稀少,达不到更新所需要数量。经过25年和40年发育的人工林,大于Ⅳ的个体数量变化不大,说明刺槐种群个体直径生长停止或者极为缓慢。

|
图 1 不同生境及年龄阶段刺槐人工林径级结构 Figure 1 The diameter-class structures of R. pseudoacacia plantations in different habitats and age stages |
同龄条件下,林木高度能反映树木对生境的适合程度。不同生境及年龄阶段刺槐人工林高度级结构(图 2)可以看出,造林25年和40年后,刺槐人工林阳坡的个体高度主要集中在Ⅳ级(8 m),分别占总个体数的49%和44%;阴坡的个体高度主要集中在Ⅴ级(10 m),占总个体数的31%和30%。总体看,刺槐人工林种群平均高度,阴坡>阳坡。造林25年和40年后高度级结构差异不大,说明造林25年以后,刺槐高度生长停止或者极为缓慢。

|
图 2 不同生境及年龄阶段刺槐人工林高度级结构 Figure 2 The height-class structures of R. pseudoacacia plantations in different habitats and age stages |
综合径级结构(图 1)和高度结构(图 2)特征,并通过树干解析加以研究确定,可以判断造林25年后,刺槐人工林基本达到数量成熟阶段,停止生长或者生长缓慢,应该进行适度间伐,以获取最大经济效益。
2.2 不同发育阶段阴坡与阳坡刺槐人工林物种重要值和多样性 2.2.1 林分特征从表 1丘陵沟壑区不同生境及年龄阶段刺槐人工林林分特征可以看到,在造林10年后,林分郁闭度基本达到较高水平,无枝条枯死; 在10 ~ 25年阶段,种群密度、盖度以及平均个体基径、冠幅、高度等参数增长很快,说明刺槐在此期间为速生期。从造林后25 ~ 40年阶段,各类参数增长幅度减缓,大部分差异不显著。种群平均个体生长发育参数在前期(25年前)增长快,主要是由于刺槐个体较小,林地内的水、肥、空间能够满足其需求。25年以后各参数增长趋缓,表明随着群落发育,种群个体对水、肥、光需求更大,不能满足生长发育所需,已经达到林地最大的承载能力(张文辉等,2004)。
|
|
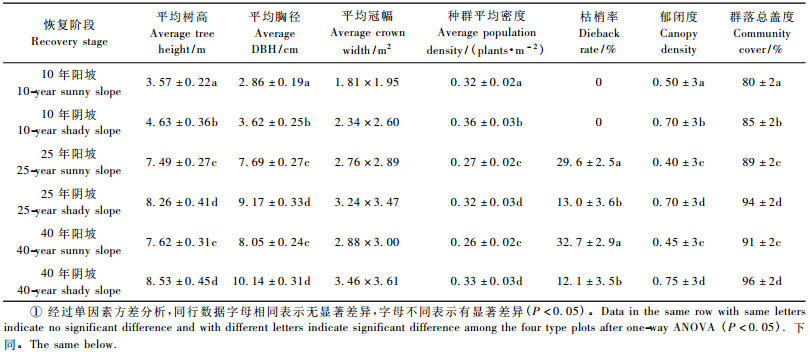
比较不同生境群落特征,阴坡明显优于阳坡。以25年刺槐样地为例,生长在阳坡的刺槐林平均高度为7.62 m,平均个体枯梢率达到29.6%,发育不良,出现小老树现象; 由于个体死亡,密度降低为27株·(10 m)-2。生长在阴坡的刺槐林平均高度为8.53 m,枯梢率只有13%,小老树现象不明显,造林后死亡量少,平均密度为33株·(10 m)-2,群落具有持续发育潜力。
2.2.2 林下物种重要值及其多样性变化人工林地不同发育时期物种重要值反映群落演化和物种替代的过程。表 2不同生境及年龄阶段刺槐人工林物种的重要值表明,造林10年后,进入群落的乡土植物,多数是1年生草本,并且能很快增加林内总盖度,提高林分防护效益。造林25年后,进入群落的乡土植物,多数是多年生草本或者灌木。乡土植物进入或者退出群落是自然发生的,对群落总盖度影响不大。
|
|
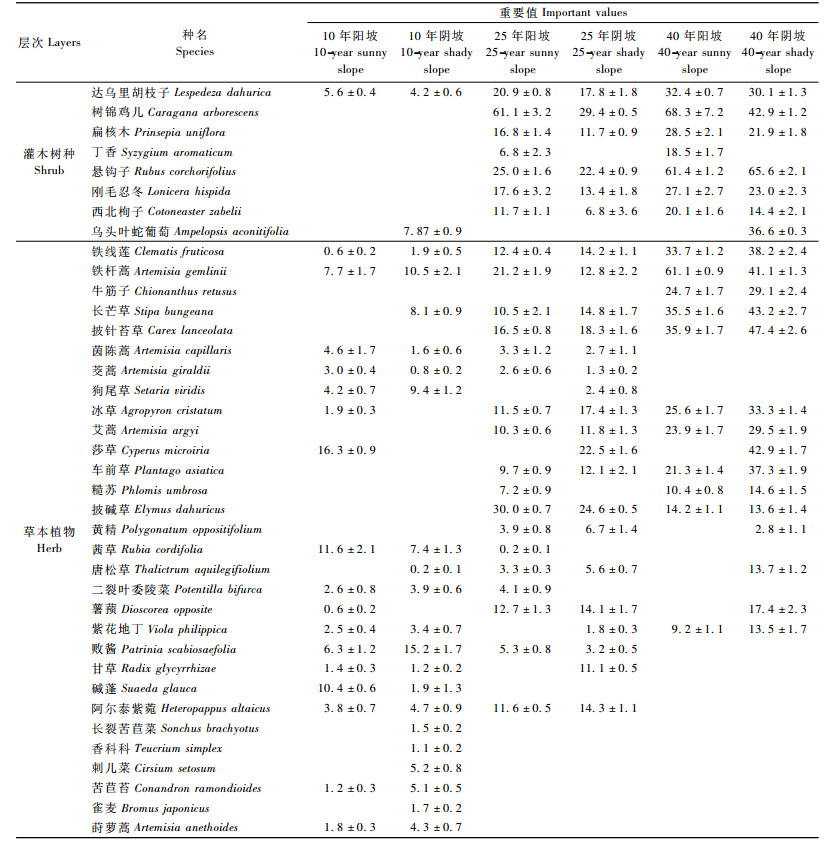
林下的灌、草植物组成差异在一定程度上反映林地生境状况。阴坡与阳坡林分中物种差异,主要是生境条件对物种的选择,如悬钩子、披针苔草(Carex lanceolata)、艾蒿、长芒草等物种在阴坡样地中分布较多; 而达乌里胡枝子、扁核木、铁杆蒿、披碱草(Elymus dahuricus)等则大多分布在阳坡样地中。无论哪种生境,刺槐人工林造林后,随着当地乡土植物种类侵入,形成具有乔、灌、草复层结构的林分,都能发挥防护功能。
表 2表明,随着发育时间的改变,灌木、草本植物组成有明显差异:同一发育阶段的群落物种组成,阴坡物种多于阳坡物种。从各物种重要值的排序结果可以看出,不同发育阶段的刺槐人工林林下植被草本植物相对都较发达,且灌-草植物中物种多为中生植物,形成相对浓密的灌木树种和草本植物,发挥着保持水土的功能,群落有持续发育潜力(张长庆等,2009; 张文辉等,2004)。表 2也展示不同发育阶段种类组成的变化情况:在发育初期刺槐人工林群落的灌木物种郁闭度较小时,阳性物种出现的频度高,随着灌木盖度增加,阳性物种减少,偏中生性的物种逐渐出现,1年生草本如碱蓬(Suaeda glauca)、茜草(Rubia cordifolia)等总是处于主要种群的地位。发育一段时间后,多年生草本如铁杆蒿、披碱草等出现,频度增加,1年生草本植物生长受到抑制,频度逐渐减少,同时达乌里胡枝子、悬钩子等灌木幼苗也开始出现。随着群落发育,灌丛在整个群落中占到相当的比例,盖度上升,一些典型喜光草本受到抑制,种类继续减少,茜草、甘草(Radix glycyrrhizae)、二裂叶委陵菜(Potentilla bifurca)的频度下降,一些偏中生性的草本植物如艾蒿、对光要求不严格的披针苔草等开始出现。在40年以后阴坡生境,刺槐林中的中性草本和灌木树种仍然占主导地位,随着群落发育,灌木树种盖度增加,养分、水分等生境将得到改善,群落内发生物种间重组,整体向中生性方向发展。
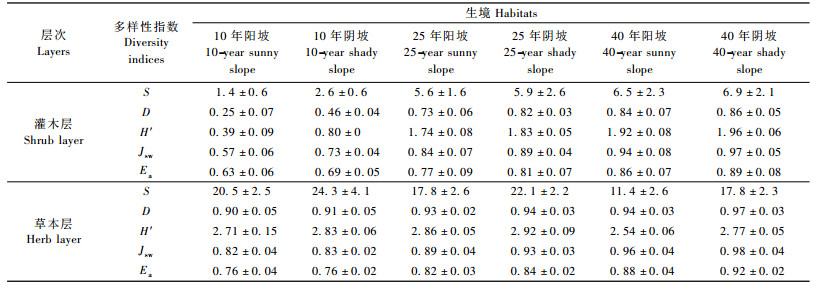
林地物种多样性是评价林地生境的重要指标。表 3是不同生境及年龄阶段刺槐人工林灌、草层的物种多样性指数。在造林10 ~ 25年阶段,丰富度指数、多样性指数和均匀度指数增加很快,说明刺槐在此期间为速生期; 灌、草层的物种多样性指数在前期(25年前)增长快,主要是由于林地内的水、肥、空间能够满足其需求。从造林后25 ~ 40年阶段,各指数增长趋缓,大部分差异不显著; 表明随着群落发育,灌、草层对水、肥、光需求更大,不能满足生长发育所需,已经达到林地最大的承载能力。比较不同生境林内物种多样性指数,生长在阴坡的刺槐人工林灌木层,其物种丰富度指数、多样性指数和均匀度指数大多较对应的阳坡高; 草本层不同发育阶段比较大致呈现出阴坡>阳坡的趋势,生长在阴坡的刺槐人工林草本层,其物种丰富度指数、多样性指数和均匀度指数大多较阳坡高; 表明在黄土高原以干旱为主的生境中,随着群落发育,灌木层盖度增加,群落丰富度、多样性、均匀度等在逐步改善。阴坡生长状况相对较好,群落具有持续发育潜力。
|
|
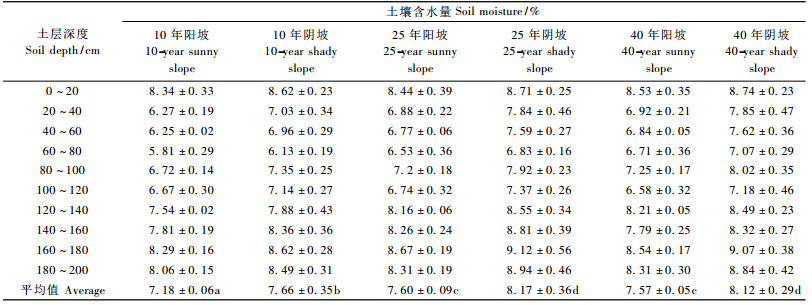
从表 4不同生境及年龄阶段刺槐人工林地0 ~ 200 cm土层水分含量特征可以看出,样地的土壤含水率均较低,不同发育阶段的阳坡林地土壤水分含量都在萎蔫系数附近(邝立刚,2001),尽管阴坡刺槐人工林发育正常,但也遭受干旱的威胁。在不同发育阶段同一生境下土壤含水率差异均不明显; 但在不同生境条件下,阴坡和阳坡土壤含水率差异显著,且阴坡的土壤含水量普遍高于阳坡。样地内100 ~ 120 cm深度都有不同程度的土壤干层(杨文治等,1992)现象出现,在这个深度阴坡的土壤含水量略高于阳坡。这些也可能是导致阳坡与阴坡刺槐林个体生长发育、种群径级结构和群落特征差异的根本原因。
|
|
林地养分反映林地生境条件。从不同生境及年龄阶段刺槐人工林地土壤0 ~ 20 cm养分特征(表 5)来看,在刺槐人工林群落发育的前25年中,土壤肥力增加明显,有机质、速效氮、有效磷、速效钾均有明显增加,在25 ~ 40年阶段增加不明显。在不同生境条件下,阴坡和阳坡土壤养分差异显著,阳坡的土壤养分含量偏低,阴坡土壤养分含量较高。这表明土壤养分水分条件与林分生长状况是相互联系的。阴坡水分条件相对较好,地上乔灌草生长旺盛,枯枝落叶量大,水保功能强,土壤水肥条件自然也就好(张婷等,2007)。这可能也是导致阳坡与阴坡刺槐林个体生长发育、种群径级结构和群落特征差异的原因。
|
|
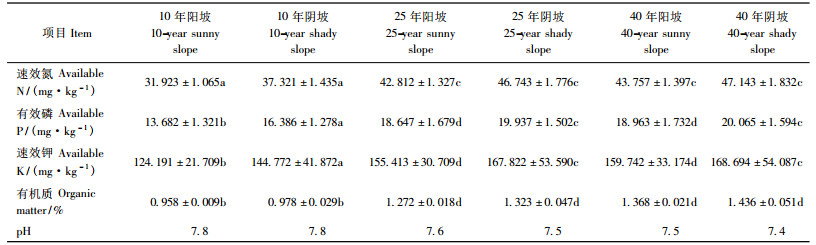
不同发育阶段刺槐人工林种群统计表明,密度、基径、冠幅、高度、多样性指数等指标等均呈现: 40年>25年>10年的趋势,但40年与25年林分差异不显著。造林后的中前期阶段,多样性增加,群落生态功能增加,对群落天然化发育有利。造林后25年阶段阴坡生境的种群萌生幼苗数量增加,种群呈现可以自然更新的趋势。从不同发育阶段和不同立地条件就可以看出,尽管阳坡刺槐人工林胸径、树高均达到最大值,并产生少量萌生幼苗,但数量相当有限,很难使刺槐人工林种群持续发育。这说明在阴坡生境,刺槐种群通过天然化培育,实现生态功能持续增强,不会出现周期性衰退现象。
3.2 人工群落发育过程中林地水分、养分和物种多样性有利于保持群落的持续发育刺槐人工林在造林经过25年发育,林木达到成熟阶段,林下物种多样性、土壤养分、水分达到比较稳定状态; 阴坡与阳坡生境的人工刺槐林比较,阴坡生境的林分,建群种在不同发育阶段,个体生长发育良好,林地物种多样性丰富,土壤养分水分相对优越,土壤干层的现象不明显,群落具有持续发育潜力。而阳坡刺槐人工林建群种个体发育不良,林分物种多样性、土壤水分养分相对较差,有明显的土壤干层现象,不具备持续发育特征。
3.3 在黄土高原丘陵沟壑区,阴坡生境通过人工林天然化培育,可以实现持续利用,而阳坡生境不适合人工林培育经过25年发育刺槐人工林种间关系已经达到较为稳定的程度,说明通过对阴坡生境刺槐人工林天然化培育,可以实现持续利用。未来管理中,应该减少人为干扰,为乡土树种的进入创造条件,以保持种间关系和谐和生态功能持续发展。在造林25年后,应通过适度间伐病虫木、劣质木,采取恰当的抚育措施提高林分质量,促进伐桩萌芽,促使种群向异龄方向发展(罗菊春,2006)。未来丘陵沟壑区应尽量避免在阳坡生境营造刺槐林,现有林地也应逐步改造,以沙棘(Hippophae rhamnoides)等耐旱灌木取代刺槐,从而实现人工群落持续发育。
曹扬, 赵忠, 渠美, 等. 2006. 刺槐根系对深层土壤水分的影响[J]. 应用生态学报, 17(5): 765-768. |
韩蕊莲, 侯庆春. 2003. 延安试区刺槐林地在不同立地条件下土壤水分变化规律[J]. 西北林学院学报, 18(1): 74-76. |
侯庆春, 韩蕊莲, 李宏平. 2000. 关于黄土丘陵典型地区植被建设中有关问题的研究: Ⅰ.土壤水分状况及植被建设区划[J]. 水土保持研究, 7(2): 102-109. |
蒋志刚, 马克平. 1997. 保护生物学[M]. 北京: 科学技术出版社: 40-41.
|
邝立刚. 2001. 旱坡地枣树的萎蔫现象[J]. 东北林业大学学报, 29(4): 101-103. |
李立平, 张佳宝, 邢维芹, 等. 2003. 土壤速效氮磷钾测定进展[J]. 土壤通报, 34(5): 483-488. |
卢金伟, 李占彬. 2002. 陕北黄土坡面水分和养分的空间变异性[J]. 山地学报, 20(1): 108-111. |
罗菊春. 2006. 抚育改造是森林生态系统经营的关键性措施[J]. 北京林业大学学报, 28(1): 121-124. |
马克平, 刘玉明. 1994. 生物群落多样性的测度方法:Ⅰ. α多样性的测度方法(下)[J]. 生物多样性, 2(4): 231-239. |
阮成江, 李代琼. 2002. 黄土丘陵区沙棘群落特性及林地水分、养分分析[J]. 应用生态学报, 13(9): 1061-1064. |
王义凤. 1991. 黄土高原地区植被资源及其合理利用[M]. 北京: 中国科学技术出版社: 23-24.
|
杨文治, 余存祖. 1992. 黄土高原区域治理与评价[M]. 北京: 科学出版社: 241-297.
|
杨文治. 2001. 黄土高原土壤水资源与植树造林[J]. 自然资源学报, 16(5): 433-438. DOI:10.11849/zrzyxb.2001.05.006 |
张长庆, 张文辉. 2009. 黄土高原不同立地条件下刺槐人工林种群的无性繁殖与更新[J]. 西北农林科技大学学报, 37(1): 135-144. |
张婷, 张文辉, 郭连金, 等. 2007. 黄土高原丘陵区不同生境小叶杨人工林物种多样性及其群落稳定性分析[J]. 西北植物学报, 27(2): 340-347. |
张文辉, 郭连金, 徐学华, 等. 2004. 黄土丘陵区狼牙刺种群恢复及群落土壤水分养分效应[J]. 水土保持学报, 18(6): 49-53. |
张文辉, 刘国斌. 2009. 黄土高原植被恢复与建设策略[J]. 中国水土保持, (1): 24-27. |
张文辉, 徐学华, 李登武, 等. 2004. 黄土高原丘陵沟壑区狼牙刺群落恢复过程中的种间联结性研究[J]. 西北植物学报, 24(6): 1018-1023. |
张仰渠. 1989. 陕西森林[M]. 北京: 中国林业出版社: 250-270.
|