 2010, Vol. 46
2010, Vol. 46文章信息
- 肖复明, 范少辉, 汪思龙, 官凤英, 于小军, 申正其
- Xiao Fuming, Fan Shaohui, Wang Silong, Guan Fengying, Yu Xiaojun, Shen Zhengqi
- 毛竹、杉木人工林生态系统碳平衡估算
- Estimation of the Carbon Balance in Moso Bamboo and Chinese Fir Plantation Ecosystem
- 林业科学, 2010, 46(11): 59-65.
- Scientia Silvae Sinicae, 2010, 46(11): 59-65.
-
文章历史
- 收稿日期:2009-09-03
- 修回日期:2010-08-14
-
作者相关文章



2. 国际竹藤网络中心 北京 100102;
3. 中国科学院会同森林生态实验站 湖南会同森林生态系统国家野外科学观测研究站 会同 418307
2. International Centre for Bamboo and Rattan Beijing 100102;
3. Huitong Experimental Station of Forest Ecosystem, Chinese Academy of Sciences Huitong National Research Station of Forest Ecosystem Huitong 418307
随着对全球气候变化的重视,近年来进行的大尺度的碳平衡研究已被广泛用来分析国家和地区、生物群落和经济区的碳平衡状况,但由于生态系统的多样性,导致这种计算结果很不一致。森林生态系统作为陆地生态系统的主体,是地球上除海洋之外最大的碳库,其有机碳贮量约为1.146 × 1012 t,占全球总碳库的46.3% (Brown,1996),因此,开展森林生态系统碳平衡动态研究,对于科学预测森林对全球碳平衡和气候变化的作用具有重要意义,并已成为国际陆地碳循环研究的热点(杨玉盛等,2007)。世界森林主要分布于热带和寒温带之间,目前国内外有关国家、区域及生态系统水平的森林碳平衡研究较多,从区域上主要集中在热带、亚热带和温带地区,森林类型上主要集中在针、阔叶林上,但由于各地气候、森林植被类型等都有很大的差别,而不同的森林类型其碳源/汇功能不尽相同(李意德等,1998; 王金叶等,2000; 方精云等,2001; 2006; 方晰等,2002; 杨智杰,2007)。随着《京都议定书》的生效实施,正确评价森林对大气CO2的固定能力,较小尺度上研究某个地区、某个林种的碳平衡特征也就显得十分迫切。
毛竹(Phyllostachys edulis)和杉木(Cunninghamia lanceolata)人工林是我国南方重要的森林资源,据不完全统计,中国现有毛竹林和杉木林总面积分别为300和1 239.1万hm2以上,分别占全世界竹林总面积20%左右和全国人工林面积的26.55%,近年种植面积有不断扩大的趋势(江泽慧,2002; 杨智杰,2007),对其碳平衡动态特征的研究,杉木林主要集中在对中心分布区域碳源/汇功能评价及其不同年龄阶段碳平衡值的计算(方晰等,2002; 肖复明,2003; 肖复明等,2007; 尉海东等,2006; 杨智杰,2007; 王兵等,2008; 魏文俊等,2008),但对竹林生态系统碳素特征研究尚少(徐秋芳等,2003; 周国模等,2004; 郭起荣等,2005; 肖复明等,2007; 2008a;2008b),现有的研究主要针对经营措施对林地土壤有机碳的影响(徐秋芳等,2003; 肖复明等,2008a; 2008b),对毛竹林生态系统碳平衡特征还未见报道。在寻找控制或减轻碳释放途径的背景下,本研究通过对毛竹和杉木生态系统碳贮量及其碳平衡特征作初步研究与探讨,从碳贮量和碳吸存角度阐明毛竹林和杉木林2种不同林分类型的碳平衡特征及其在全球碳平衡中的作用,以期为杉木、毛竹次生林、低产林的改造及高效率碳吸存经营措施提供基础数据,也为正确评价不同类型的森林在全球碳平衡中的作用提供理论依据,进而为制定森林发展规划和环境保护政策提供理论基础。
1 试验地概况研究样地位于湖南省会同县境内,气候属亚热带湿润气候,年均温度16.5 ℃,极端最高温和最低温分别为36.4和-4.4 ℃,年均降雨量为1 200 ~ 1 400 mm,年均相对湿度在80%以上,年均日照1 445.4 h,无霜期304天,海拔高为300 ~ 500 m,土壤为红黄壤。毛竹林位于湖南省会同县肖家乡坡脚村(109° 53′ E,27°03′ N),试验林原为荒芜低产毛竹林,1988年冬经低产林改造后而建立的,但仍然有大小年之分,一般偶数年为大年,奇数年为小年,林分现有立竹度为1 800 ~2 400株·hm-2,林地基本没有灌木,林下植被生长季节以马唐(Digitaria sanguinalis)、鸡屎藤(Paederia scandens)、鱼腥草(Houttuynia cordata)、蛇葡萄(Ampelopsis aconitifolia)、莎草(Cyperus difformis)、铁芒萁(Dicranopteris linearis)等杂草为主。
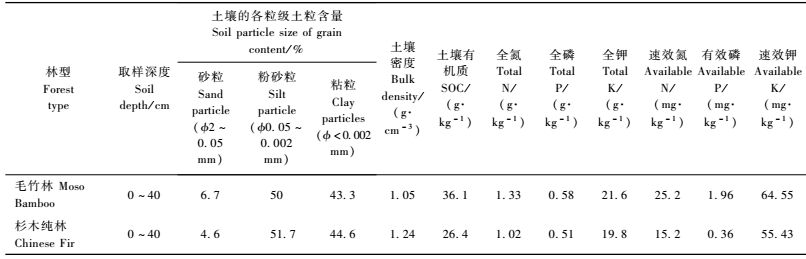
试验同时设立杉木人工林为对照,杉木人工林样地设在中国科学院会同森林生态实验站(109°30′ E,26°48′ N),海拔200 ~ 500 m,为低山丘陵地貌类型,林龄为15年生,林分现有密度为1 530株·hm-2,林分平均胸径为14.8 cm,平均树高为12.6 m,造林地前茬为杉木林,林地灌木主要以杜茎山(Maesa japonica)、格药柃(Eurya muricata),草本以狗脊(Woodwardia japonica)、中华里白(Hicriopteris chinensis)等为优势种的生态系统。2林地土壤的主要物理、化学性质如表 1所示。
|
|
毛竹生物量是2005年11月在3个20 m × 30 m标准地内进行每株立竹调查的基础上,计算出每度竹的平均立竹作为标准竹,并分别砍伐Ⅰ,Ⅱ,Ⅲ,Ⅳ度竹的标准竹各2株,分别测定竹杆、竹枝和竹叶鲜质量,并对地下部分用挖掘法每个样地挖掘4个1 m × 1 m样方,挖出竹蔸、鞭根,挖至无根为止,用水仔细漂洗,再用筛子在水中筛后捡出所有的根,风干表面水称鲜质量; 各组分取样后带回实验室,在60 ℃烘干至恒质量,并取小样105 ℃烘干,求算生物量。
毛竹年生产力的测定,根据所伐标准竹得到有关经验方程,计算出毛竹地上部分年净生产量(陈辉等,1998),公式如下:
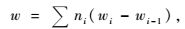
|
式中,w为毛竹地上部分年净生产量; ni为各龄竹株数; i为竹龄; wi为第i龄竹地上部干质量平均值; wi-1为第i-1龄竹的地上部干质量平均值。
地下年净生产量的测定,由标准竹得到的经验方程中毛竹地下部与地上部现存量之比率乘以地上部分年净生产量得出地下部分年净生产量,加上退笋和笋箨的年产量,再加上通过收集试验得到的年凋落物及林下植物的现存量(因每年垦挖,故以现存量推算),即可得出毛竹林的年净生产量。
杉木生物量测定采用中国科学院会同森林生态试验站现有杉木生长方程和样地调查相结合(陈楚莹等,2000)。2005年和2006年11月对3个20 m × 20 m杉木林标准地内的杉木胸高直径和树高进行每木检尺后,利用现有的杉木生长方程,计算出杉木生物量,并以年平均生物量增长量作为净生产量的估算指标。
2.2 林下植被生物量和净生产量测定2005年11月在每个标准地内分上、中、下坡位设立1 m × 1 m小样方5个,采用全挖法实测生物量。即收集毛竹林每个小样方中全部灌木、杂草和杉木林每个小样方中草本植物,混合5个小样方中的灌木、杂草,烘干至恒质量再估算干质量大小,同时用作分析样品。杉木多年生灌木生物量采用中国科学院会同森林生态试验站生物监测数据进行计算。
2.3 凋落物现存量和净生产量测定2005年11月在每个样地内各设置5个1 m ×1 m收集器,将收集器下小样方的全部枯落物收集,计算凋落物现存量; 2005年12月—2006年12月每月收集收集器中凋落物1次,按枝、叶、箨、果等组分收集凋落物,并分别称其湿质量后,在80 ℃条件下烘干后称质量,同时选取一定量的凋落物作为分析样品。
2.4 细根生物量测定细根生物量、年生长量及分解量等调查采用“连续钻取土芯法”和改进的最大值、最小值法计算模型,具体测定方法和数据见范少辉等(2009a)报道。
2.5 样品采集及化学分析方法在测定生物量的同时,按层分组采集标准株的分析样品。竹秆和杉木树干每2 m取10 ~ 20 g鲜样称量质量; 枝、叶分层(上、中、下),根系分级(按<0.2 cm,0.2 ~ 0.5 cm,> 0.5 cm,根头),土壤分层(0 ~ 20,20 ~ 40,40 ~ 60 cm)然后各层(级)分别采集分析样品,土壤各层取样分上、中、下坡3个取样点,并在每个取样点不同位置取3个点的混合样,取样点的分布基本采用S形,同时采用环刀法取原状土,测定土壤密度(刘光崧,1996)。
植物、土壤样品中碳素含量测定采用重铬酸钾法(刘光崧,1996)。
2.6 碳贮量的计算不同器官生物量与其碳含量的乘积为不同器官的碳贮量; 各器官碳贮量之和为乔木层碳贮量; 植被各组分年净生产量与其碳含量的乘积为各组分年有机碳固定量; 乔木层、灌木层、草本层、枯落物层和土壤层有机碳贮量之和为生态系统中的碳贮量。其中,土壤碳贮量计算公式为: Ss = Sd × D × S × Cc式中,Ss为土壤碳贮量(t·hm-2); Sd为土壤密度(g·cm-3); D为采样深度(cm); S为面积(hm2); Cc为土壤有机碳含量(%)。
2.7 土壤呼吸测定在毛竹和杉木各3个固定样地的每个样地内选择6个不同的位置(上、中、下坡位,每个坡位2个不同位置),采用壕沟法进行切断根系去除凋落物(- R-L)、保留根系保留凋落物(+ R + L)和保留根系去除凋落物(+ R-L) 3种处理进行试验(Hanson et al., 2000; Burton et al., 2003; 杨金艳等,2006),具体操作是2005年10月在选定的60 cm ×60 cm小样方,四周插入4块60 cm × 60 cm的硬塑料板,即插入深度为60 cm进行挖壕断根处理(壕沟深度达基岩或根系分布层以下)。自2006年1月—12月,每月中旬选择1个晴天昼夜利用CID公司的土壤呼吸室与CID-301光合测定仪进行土壤呼吸观测,每次测定前1天对小样方进行检查和测定前准备,测定时间间隔为2 h,每个处理每次仪器测定3 ~ 5 min,仪器采样速率为30 s,气室内CO2浓度降至等于大气中浓度后才能进行下一个测定。测定林地土壤呼吸的同时,采用数字式瞬时温度计(Longstem thermoneter 6310)和湿度计(HH2 moisturemeter)测定观测点附近地表温度和土壤含水量以及土层5,10,20 cm深的温度。具体测定方法和数据分析见范少辉等(2009b)报道。
2.8 毛竹林生态系统碳收支计算公式毛竹林生态系统的植被固定CO2的计算公式为:
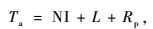
|
式中: Ta为毛竹林生态系统群落生产同化CO2总量,也是系统CO2总收入量; NI为植被年固定CO2净增量; L为年凋落物层中贮存的CO2转化量; Rp为植被年呼吸释放CO2量。
毛竹林生态系统CO2支出的计算公式为:
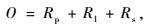
|
式中,O为毛竹林生态系统的CO2支出总量; Rp为植被年呼吸释放的CO2量; Rl为年凋落物分解释放的CO2量; Rs为土壤呼吸所释放的CO2量。
整个毛竹林生态系统的CO2收支公式为:
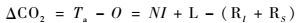
|
式中,ΔCO2为整个生态系统CO2的平衡值,即生态系统的净生产量(NEP)。
3 结果与分析 3.1 生态系统碳贮量及其分配格局由表 2可知:毛竹林生态系统碳贮量为144.3 t·hm-2,其中,土壤层碳贮量最大,为110.95 t·hm-2,占76.89%,其次是毛竹层为31.97t·hm-2,占22.16%,林下植被和凋落物层分别为0.64和0.74 t·hm-2,分别占0.44%和0.51%,毛竹层与土壤层之比为1: 3.47,说明林地碳贮量是1个极大的碳贮量库。15年生杉木人工林生态系统中碳贮量为152.52 t·hm-2,其中也是土壤层最大,为95.18 t·hm-2,占总贮量的62.40%,其次为乔木层,占总贮量的34.66%,林下植被层和凋落物层分别只占0.74%和2.24%,乔木层与土壤层碳贮量之比为1: 1.80。
|
|
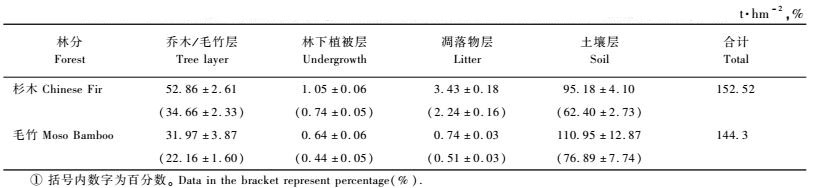
森林生态系统的生产力研究的主要内容之一是要确定系统同化CO2的能力。由表 2和3可知:15年生杉木人工林植被层贮存碳总量为53.91 t·hm-2,相当于同化CO2的量为197.67 t·hm-2,年固定有机碳量为7.36t·hm-2 a-1,相当于每年同化CO2的量为27.04t·hm-2 a-1。毛竹林植被层碳贮量为32.71 t·hm-2,只占杉木林的60.67%,但是,毛竹林是异龄林分,通常采取择伐作业,隔年伐去Ⅳ度以上竹。因此从生态系统的角度来看,毛竹林分永远处于生长动态平衡之中,并可以近似认为每次采去毛竹的生物量相当于现存生物的1 /4,以此换算,毛竹林砍伐掉63.94 t·hm-2,约为杉木林现存量的1.21倍。由表 3可知:毛竹林年固定有机碳量为10.60t·hm-2 a-1,相当于年同化CO2量38.87 t·hm-2 a-1,是杉木林的1.44倍。毛竹和杉木细根年死亡量分别占年凋落物总量的25.8%和9.7%,说明毛竹林细根的归还量是毛竹林群落生产力的重要组成部分之一。
|
|
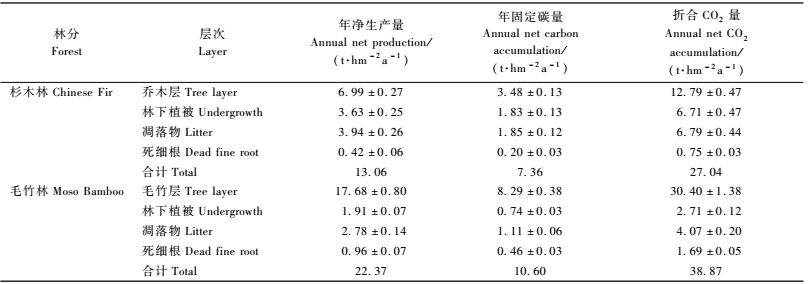
由表 4可知:毛竹林地土壤年释放CO2量为33.941 4 t·hm-2,月平均为2.828 4 t·hm-2,其中,林地土壤和凋落物分别占87.8%和12.2%。杉木林地土壤年释放CO2量为19.879 7 t·hm-2,月平均为1.656 4 t·hm-2,其中,林地土壤和凋落物分别占75.6%和24.4%。在一年12个月中,不论是毛竹林还是杉木林林地土壤释放CO2量的变化趋势基本一致,最高峰值出现在7,8月份,7月两者分别为5.370 7和3.090 5 t·hm-2,8月份分别为6.775 4和2.970 6 t·hm-2,最低值出现在1,2月份,分别为0.962 3和0.684 t·hm-2,说明7,8月份的高温高湿条件有利于土壤微生物和根系的生命活动,而低温干燥容易抑制他们的生命活动。
|
|
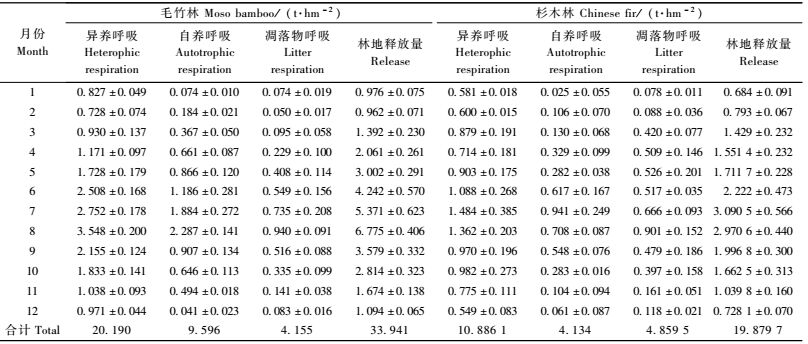
2个群落土壤CO2释放量的变化趋势有差别,即毛竹林土壤呼吸速率随时间的波动比杉木林剧烈。这种波动可以通过其最大值与最小值的比来反映,毛竹林的最大值/最小值比为7.04,而杉木林则为4.34,相差较大,这也从另外一侧面说明毛竹林的土壤CO2释放更容易受到外界气候因子的影响。
3.4 生态系统碳收支计算森林生态系统碳平衡包括输入与输出2个过程,输入与输出的差值即为生态系统的净生产量(NEP),若NEP为正,表明生态系统是CO2的汇,反之则为CO2源。碳的输入主要是植被对CO2的固定,输出包括群落呼吸、土壤呼吸和凋落物分解释放CO2。
根据以上的研究,参照文献(李意德等,1998; 方晰等,2002; 杨玉盛等,2007)的统计方法,计算毛竹和杉木林生态系统的CO2固定吸收值及生态系统各分量对大气的CO2排放过程等一系列参数(表 5),从表 5毛竹林年固定CO2总量为38.87t·hm-2 a-1,林地土壤异养呼吸和凋落物每年以呼吸形式排放CO2的量分别为20.19和4.16t·hm-2 a-1,整个毛竹林生态系统年净固定CO2的量为14.52 t·hm-2 a-1,折合成净碳量为3.96t·hm-2 a-1; 杉木林年固定CO2总量为26.95t·hm-2 a-1,林地土壤和凋落物每年以呼吸的形式排放CO2的量分别为10.89和4.86 t·hm-2 a-1,整个杉木林生态系统年净固定CO2的量为11.21t·hm-2 a-1,折合成净碳量为3.07 t·hm-2 a-1。
|
|

1) 毛竹林植被年固定CO2总量为38.87t·hm-2 a-1,是杉木林的1.44倍,高于尖峰岭热带山地雨林(31.59 t·hm-2 a-1) (李意德等,1998)、亚热带35年生青冈(Cyclobalanopsis glauca)常绿阔叶林(28.43 t·hm-2 a-1) (李铭红等,1996),但比我国常绿阔叶林(45.06 t· hm-2 a-1)和福建格氏栲(Castanopsis kawakamii)和木荷(Schima superba)林(50.01和44.15 t·hm-2 a-1)低(周玉荣等,2000; 何宗明等,2003; 杨智杰,2007; 杨玉盛等,2007)。杉木人工林高于同一试验区的10年和11年生杉木人工林(20.17和22.30 t·hm-2 a-1),但低于福建36年生杉木林(24.20 t·hm-2 a-1)和33年生福建柏(Fokienia hodginsii)林(36.33 t·hm-2 a-1) (方晰等,2002; 何宗明等,2003; 杨玉盛等,2007),说明森林植被年固定CO2量高,不仅与树种有关,而且还与其林龄、立地等条件有关,因此,加强对不同区域、不同类型森林植被固碳能力的研究将有助于正确评价不同森林类型的固碳能力。
2) 毛竹林年净碳汇为3.96 t·hm-2 a-1,是杉木林的1.29倍,与澳大利亚北部热带萨瓦纳森林(3.8 t·hm-2 a-1)及福建36年生杉木林(3.615 t·hm-2 a-1)相近(Chen et al., 2003; 杨玉盛等,2007),高于亚马逊热带森林(1.00 t·hm-2 a-1)、温带落叶混交林(1.4 ~ 2.8 t·hm-2 a-1),尖峰岭热带山地原始林(0.373 t·hm-2 a-1),但低于我国常绿阔叶林(7.28 t·hm-2 a-1)、落叶阔叶林平均水平(4.07 t· hm-2 a-1)、福建36年生格氏栲林(7.656 t·hm-2 a-1) (Grace et al., 1995; 杨玉盛等,2007; 周玉荣等,2000)。造成这种差异性可能不仅与树种生物学特性有关,可能还与毛竹和杉木林立地条件的差异有关。此外,目前有关碳汇研究的统计方法不一致也可能是导致相关文献结果有差异的原因,如方晰等(2002)研究表明:会同10年生杉木林表现为净碳源(- 0.427 t·hm-2 a-1),11年生杉木才表现为碳汇(0.168 t·hm-2 a-1),但其在碳平衡计算中直接利用土壤呼吸代替异养呼吸; 方精云等(2006)研究白桦林、辽东栎林和油松林的净碳汇(NEP)分别为0.95,-0.29和4.08 t·hm-2 a-1,但其在碳平衡计算时NPP组成中忽略地下净生产力部分。因此,采用统一规范的研究方法,获取大量有代表性的实测数据,是减少区域尺度碳平衡研究中的不确定性,从而可更精确地评价不同森林类型在全球变化中的作用,也为今后营造碳汇林树种选择提供科学依据。
3) 毛竹林碳平衡中2个分量的比率,即土壤异养呼吸量/乔木层净碳吸存量为0.54,是杉木林的1.31倍,也就是说集约经营毛竹林植被净初级生产力(NPP)和土壤CO2排放量同时增加的现象。因此,开展毛竹林生态系统碳库的循环周期与库存功能研究,建立毛竹林生态系统碳循环动力学模型,分析毛竹林在中国森林碳平衡中的作用和增汇潜力,进而提出毛竹林生态系统碳平衡调控技术措施,是今后的研究方向。
陈楚莹, 廖利平, 汪思龙. 2000. 杉木人工林生态学[M]. 北京: 科学出版社.
|
陈辉, 洪伟, 兰斌, 等. 1998. 闽北毛竹生物量与生产力的研究[J]. 林业科学, 34(sp1): 60-64. |
范少辉, 肖复明, 汪思龙, 等. 2009a. 毛竹林细根生物量及其周转[J]. 林业科学, 45(7): 1-6. |
范少辉, 肖复明, 汪思龙, 等. 2009b. 湖南会同林区毛竹林地的土壤呼吸[J]. 生态学报, 29(11): 5971-5977. |
方精云, 陈安平. 2001. 中国森林植被碳库动态变化及其意义[J]. 植物学报, 43(9): 967-973. |
方精云, 刘国华, 朱彪, 等. 2006. 北京东灵山三种温带森林生态系统的碳循环[J]. 中国科学D辑:地球科学, 36(6): 533-543. |
方晰, 田大伦, 项文化, 等. 2002. 第二代杉木中幼林生态系统碳动态与平衡[J]. 中南林学院学报, 22(1): 1-6. |
郭起荣, 杨光耀, 杜天真, 等. 2005. 中国竹林的碳素特征[J]. 世界竹藤通讯, 3(3): 25-28. |
何宗明, 李丽红, 王义祥, 等. 2003. 33年生福建柏人工林碳库与碳吸存研究[J]. 山地学报, 21(3): 298-303. |
江泽慧. 2002. 世界竹藤[M]. 沈阳: 辽宁科学技术出版社.
|
李铭红, 于明坚, 陈启瑺, 等. 1996. 青冈常绿阔叶林的碳素动态[J]. 生态学报, 16(6): 645-651. |
李意德, 吴仲民, 曾庆波, 等. 1998. 尖峰岭热带山地雨林生态系统碳平衡的初步研究[J]. 生态学报, 18(4): 371-378. |
刘光崧. 1996. 中国生态系统研究网络观测与分析标准方法———土壤理化分析与剖面描述[M]. 北京: 中国标准出版社.
|
王兵, 魏文俊, 李少宁, 等. 2008. 中国杉木林生态系统碳储量研究[J]. 中山大学学报, 47(2): 93-98. |
王金叶, 车克钧, 蒋志荣. 2000. 祁连山青海云杉林碳平衡研究[J]. 西北林学院学报, 15(1): 9-14. |
尉海东, 马祥庆. 2006. 中亚热带不同发育阶段杉木人工林生态系统碳贮量研究[J]. 江西农业大学学报, 28(2): 239-243. |
魏文俊, 王兵, 白秀兰. 2008. 杉木人工林碳密度特征与分配规律研究[J]. 江西农业大学学报, 30(1): 73-80. |
肖复明, 范少辉, 汪思龙, 等. 2007. 毛竹?杉木人工林生态系统碳贮量及其分配特征[J]. 生态学报, 27(7): 2794-2801. |
肖复明, 范少辉, 汪思龙, 等. 2008a. 毛竹林地土壤团聚体稳定性及其对碳贮量影响研究[J]. 水土保持学报, 22(2): 131-134. |
肖复明, 范少辉, 汪思龙, 等. 2008b. 毛竹林土壤有机碳及微生物量碳特征研究[J]. 水土保持学报, 22(6): 128-131. |
肖复明. 2003. 杉木人工林生态系统碳平衡的研究. 江西农业大学硕士学位论文.
|
徐秋芳, 徐建明, 姜培坤. 2003. 集约经营毛竹林土壤活性有机碳库研究[J]. 水土保持学报, 17(4): 15-21. |
杨金艳, 王传宽. 2006. 东北东部森林生态系统土壤呼吸组分的分离量化[J]. 生态学报, 26(6): 1640-1647. |
杨玉盛, 孙光水, 王义祥, 等. 2007. 格氏栲人工林和杉木人工林碳吸存与碳平衡[J]. 林业科学, 43(3): 113-117. |
杨智杰. 2007. 杉木?木荷人工林碳吸存与碳平衡研究. 福建农林大学硕士学位论文.
|
周国模, 姜培坤. 2004. 毛竹林的碳密度和碳贮量及其空间分布[J]. 林业科学, 40(6): 20-24. DOI:10.11707/j.1001-7488.20040604 |
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳库和碳平衡[J]. 植物生态学报, 24(5): 518-522. |
Brown S. 1996. Present and potential roles of forests in the global climate change debate[J]. Unasylva, 47: 3-10. |
Burton A J, Pregitzer K S. 2003. Field measurement of root respiration indicate little to no seasonal temperate acclimation for sugar maple and red pine[J]. Tree Physiology, 23: 273-280. DOI:10.1093/treephys/23.4.273 |
Chen X Y, Hutley L B, Eamus D. 2003. Carbon balance of a tropical savanna of northern Australia[J]. Oecologia, 137: 405-416. DOI:10.1007/s00442-003-1358-5 |
Grace J, Lloyd J, Miranda A C, et al. 1995. Carbon dioxide uptake by an undisturbed tropical rain forest in South-West Amazonia 1992-1993[J]. Science, 270: 778-780. DOI:10.1126/science.270.5237.778 |
Hanson P J, Edwards N T, Garten C T, et al. 2000. Separating root and soil microbial contributions to soil respiration: A review of methods and observations[J]. Biogeochemistry, 48: 115-146. DOI:10.1023/A:1006244819642 |