 2010, Vol. 46
2010, Vol. 46文章信息
- 顾地周, 李玉梅, 姜云天
- Gu Dizhou, Li Yumei, Jiang Yuntian
- 水曲柳嫩茎尖直接再生芽苗及其离体保存的激素组合筛选
- Selection of Proper Hormone Combinations for Direct in vitro Plant Regeneration from Bud Tips and Their in vitro Preservation for Fraxinus mandshurica
- 林业科学, 2010, 46(10): 178-182.
- Scientia Silvae Sinicae, 2010, 46(10): 178-182.
-
文章历史
- 收稿日期:2009-08-06
- 修回日期:2010-06-01
-
作者相关文章


2. 吉林师范大学生态环境研究所 四平 136000
2. Institute of Ecological Environment, Jilin Normal University Siping 136000
水曲柳(Fraxinus mandshurica)属于木犀科(Oleaceae)白蜡树属(Fraxinus),是我国东北地区重要的珍贵硬阔树种之一,国家二级重点保护植物(汪松等,2004)。主要分布于小兴安岭和长白山地区,在俄罗斯远东地区、日本亦有分布。水曲柳与胡桃楸(Juglans mandshurica)、黄菠萝(Phellodendron amurense)被称为我国东北珍贵的“三大硬阔树种”,素有“东北木材三宝”的美称。水曲柳树形优美,树冠宽大,而且抗性较强,具有较高的景观价值,其树皮入药,中药称“秦皮”,具有较高的药用价值。小兴安岭和长白山林区作为水曲柳的中心分布区和主要生产基地之一,由于开发历史较早,加之长期的过度采伐和不合理利用,目前已经难以见到优质大径用材。
为了更好、更合理地开发利用水曲柳这一珍贵树种资源,在保护现有野生水曲柳资源的同时,可大力营造人工水曲柳林。目前,水曲柳主要靠种子繁殖,但对于筛选出的优良品系来说,通过建立种子园,生产种子应用于育苗造林,需时太长。无性繁殖是优良品系资源直接利用的优良途径,但是其中的扦插和嫁接途径本身繁殖系数较低,而且对于水曲柳来说,这些方法难度更大,在生产上应用不现实(沈海龙等,2005)。而同属于无性繁殖体系的组织培养方法,却具有繁殖系数大、繁殖速度快的特点。水曲柳组织培养研究仅见少量报道,主要集中在未成熟胚的离体培养(张惠君等,2003)、下胚轴不定芽诱导(谭燕双等,2003)、腋芽培养(张丽杰等,2007)、体细胞胚与合子胚发生的细胞学研究(孔冬梅等,2006)等方面,这些研究结果都比较初步,尚未达到实际生产性应用的水平。本试验应用均匀设计法对水曲柳嫩茎尖直接再生芽苗、芽苗生根及试管苗保存的培养基进行筛选,旨在筛选适宜的水曲柳嫩茎尖组织培养及其种质试管保存的培养方案,以便为水曲柳的离体快繁及工厂化育苗开辟一条适宜的途径。
1 材料与方法 1.1 水曲柳外植体材料的处理2005年3月,于吉林省通化白鸡腰国家级自然保护区采水曲柳休眠枝,在实验室内水培促使腋芽萌发。待休眠芽萌发并长至1.0 cm时,将茎尖剪下,在超净工作台上用75%酒精涮洗15 s,再用3%的次氯酸钠溶液浸泡10 min,然后无菌水冲洗6次。用无菌滤纸吸干表面水分后,切除灭菌剂损伤的部分后留3~5 mm作为外植体备用。
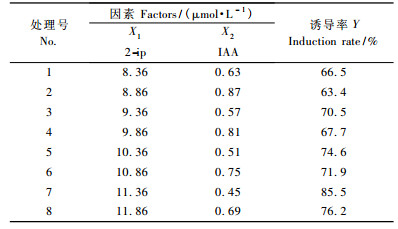
1.2 水曲柳嫩茎尖直接再生芽苗诱导培养的激素组合的筛选以N6(由对比试验可知N6较其他常用培养基诱导的芽苗整齐且速度快)为基本培养基(曹孜义等,2002),添加蔗糖20 g·L-1,琼脂粉7.0 g·L-1,再附加不同质量浓度的细胞分裂素2-ip (异戊烯基腺嘌呤) (由预试验确定为8. 36~11.86 μmol·L-1)和生长素IAA (由预试验确定为0.45~0.87 μmol·L-1),高压灭菌前调节pH为6.0,外植体在温度(24 ± 2) ℃、光照强度21.6 μmol ·m-2 s-1、光照周期每天14 h条件下培养。为提高水曲柳再生芽苗的速度和诱导率,采用均匀设计法(顾地周等,2008a; 2009a),每个处理接种茎尖数为10个,重复3次,选用U8(82)均匀表,考察2-ip和IAA不同质量浓度交叉配比对诱导率的影响。嫩茎尖培养40天统计诱导率,筛选最适合水曲柳嫩茎尖基部芽苗的诱导培养的激素组合。
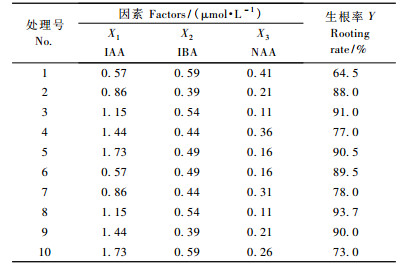
1.3 水曲柳再生芽苗生根培养的激素组合的筛选以1/4N6为基本培养基(由对比试验可知1/4N6较其他培养基生的根粗壮),加入蔗糖10 g·L-1,琼脂7.0 g·L-1,并附加不同浓度的IAA(由预试验确定为0.57~1.73 μmol·L-1)、IBA (由预试验确定为0.39~0.59 μmol·L-1)和NAA (由预试验确定为0.11~0.41 μmol·L-1),高压灭菌前调节pH为6.0,将2.0~3.0 cm的组培苗在温度(24 ± 2) ℃、光照强度18 μmol·m-2s-1、光照周期每天12 h条件下培养。为了提高水曲柳组培苗的生根率,选用U10(103)均匀表,每个处理接种茎段数为10个,重复3次,考察生长素IAA,IBA和NAA不同质量浓度交叉配比对生根率的影响。组培苗培养35天统计生根率,筛选最适合水曲柳组培苗生根培养的激素组合。
1.4 水曲柳单芽试管保存培养的激素组合的筛选采用“激素超量”的方法,即通过加入过量的细胞分裂素2-ip抑制单芽基部芽苗分化,并促使缓慢生长的方法在试管内保存水曲柳种质。以N6为基本培养基,加入蔗糖20 g·L-1,并附加不同质量浓度的2-ip (由预试验确定为12.30~15.80 μmol·L-1)和KT(由预试验确定为5.11~8.33 μmol·L-1)。在温度为(22 ± 2) ℃,光照强度18 μmol·m-2 s-1,光照周期每天8 h条件下对水曲柳单芽进行试管保存。为了降低芽苗分化率和单芽生长速度,选用U8(82)均匀表,每个处理数为10个,重复3次,考察不同质量浓度的2-ip和KT交叉配比对水曲柳单芽分化及生长的影响,保存12个月统计分化率,筛选最合适的试管保存培养的激素组合。
1.5 数据处理与分析采用均匀设计法进行初步规律设计性试验,数据分析处理应用均匀设计软件(Uniform Design 3.0V)。
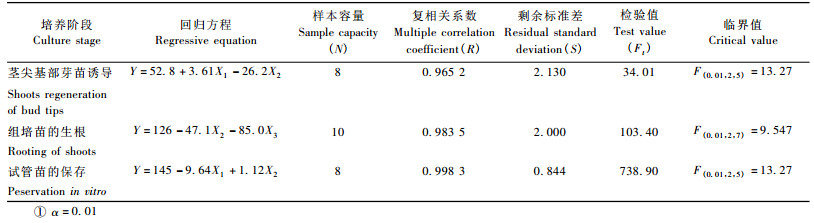
2 结果与分析 2.1 N6培养基中不同质量浓度2-ip和IAA配比对水曲柳嫩茎尖基部直接再生芽苗的影响N6培养基中不同质量浓度2-ip和IAA配比对水曲柳嫩茎尖直接再生芽苗影响的试验所得数据(表 1)经均匀设计软件分析处理后(表 2)可知: Ft > F(0.01,2,5),表明回归方程有意义。对各方程项进行显著性检验可知:各方程项对Y影响均显著。根据回归方程求出Y的最优组合为: X1 = 11.86,X2 = 0.45,在此组合基础上求得最优解: y = 83.9,此解为回归方程的解析解,需按公式Y = y ± uα·s(其中uα为正态分布的双侧分位数,s为剩余标准差)计算出优化值区间估计为Y = 83.9 ± 8.59,即75.31%~92.49%。因2-ip超过11.86 μmol·L-1时芽苗诱导率降低。通过计算各方程项对回归的贡献值和贡献率分别为: U(1) = 126,U(1)/U = 40.8%; U(2) = 95.1,U(2)/U = 30.8%,说明2-ip对芽苗诱导率Y的贡献大于IAA。
|
|
|
|
为了避免2-ip在11. 36~11. 86 μmol·L-1之间有较高芽苗诱导率的峰值,又以2-ip为11.36,11.41,11.46,11.51,11.56,11.61,11.66,11.71,11.76,11.81和11.86 μmol · L-1,IAA 0.45 μmol·L-1做了11个处理的试验,发现2-ip在11.46 μmol·L-1时芽苗诱导率最高且诱导速度快。因此,将水曲柳嫩茎尖接种到附加2-ip 11.46 μmol· L-1和IAA 0.45 μmol·L-1的N6培养基上进行芽苗诱导培养的验证试验,重复3次,培养14天,基部产生少量颗粒状突起; 继续培养至25天,大量颗粒状突起聚集成团,培养至38天,部分突起变为锥形突起,并逐渐伸长为芽苗形成丛生芽团(图 1a),芽苗诱导率达91. 8%以上,在估计区间内,且比表 1中8个处理的诱导率均高。因此,水曲柳嫩茎尖直接再生芽苗的适宜诱导培养方案为: N6 + 2-ip 11.46 μmol·L-1 + IAA 0. 45 μmol·L-1。

|
图 1 水曲柳离体培养各培养阶段的培养物形态 Figure 1 The morphogenesis of cultures in vitro culture in various stages of F. mandshurica a.嫩茎尖基部直接再生芽苗的诱导培养 Shoots regeneration from bud tips; b.生根培养 Cultivated for rooting; c.种质试管保存 Germplasm preservation in vitro; d.解除保存后生长情况 Growth situation released from preservation. |
1/4N6培养基中不同质量浓度生长素配比对水曲柳组培苗生根影响的试验所得数据(表 3)经均匀设计软件分析处理后(表 2)可知,Ft > F(0.01,2,7),回归方程显著。对各方程项进行显著性检验可知: X2和X3方程项对Y影响均显著。根据回归方程求出Y的最优组合为: X2 = 0.39,X3 = 0.11,在此组合基础上求得最优解: y = 98.4,此解为回归方程的解析解,按公式Y = y ± uα·s计算出优化值区间估计为Y = 98.4 (± 6.98),即91.42%~105.38%。通过计算各方程项对回归的贡献值和贡献率(按偏回归平方和降序排列)分别为: U(3) = 692,U(3)/U = 84.0%; U(2) = 111,U(2)/U = 13. 4%,说明NAA对组培苗生根率Y的贡献大于IBA,又因IBA和NAA均与生根率呈负相关,猜测IBA低于0.39 μmol·L-1和NAA低于0.11 μmol·L-1时有较高的生根率,又以IBA为0,0.04,0.09,0.14,0.19,0.24,0.29,0.34和0.39 μmol·L-1,NAA为0,0.05和0.10 μmol·L-1做了9个处理的试验,发现IBA在0.29 μmol·L-1和NAA 0.10 μmol·L-1时组培苗生根率最高。因此,当水曲柳嫩茎尖诱导产生的芽苗长至2.0~3.0 cm时,将其切下接种到附加IBA 0.29 μmol·L-1和NAA 0.10 μmol·L-1的1/4N6培养基上进行生根培养的验证试验,重复3次,培养10天发现茎段基部切口处有3~5个独立的锥形颗粒出现,继续培养至20天,锥形状颗粒逐渐伸长为肉质的不定根(图 1b),培养至30天,不定根可生长至6.00 cm以上,生根率达99.5%。在估计区间内,且比表 3所列10个处理的生根率均高。可见,水曲柳组培苗的适宜生根培养方案为: 1/4N6 + IBA 0.29 μmol·L-1 + NAA 0.10 μmol·L-1。
|
|
N6培养基中不同质量浓度2-ip和KT配比对水曲柳单芽试管保存的影响实验所得数据(表 4)经均匀设计软件分析处理后(表 2)可知,Ft > F(0.01,2,5),回归方程显著。对各方程项进行显著性检验可知:各方程项对Y影响均显著。根据回归方程和研究目的(反向优化)求出Y的最优组合为: X1 = 15.80,X2 = 5.11,在此组合基础上求得最优解: y =-1.50,即视为y = 0.00,此解为回归方程的解析解,按公式Y = y ± uα·s计算出优化值区间估计为Y = 0 ± 3.40,即0~1. 90%。通过计算各方程项对回归的贡献值和贡献率(按偏回归平方和降序排列)分别为: U(1) = 895,U(1)/U = 85.1%; U(2) = 11.9,U(2)/U = 1.13%,说明2-ip对单芽基部芽苗分化率Y的贡献远远大于KT,又因KT反向优化可视为与分化率呈负相关,猜测KT低于5.11 μmol·L-1时有更低的分化率,又以KT为0,0.11,0.61,1.11,1.61,2.11,2.61,3.11,3.61,4.11,4.61和5.11 μmol · L-1,2-ip为15.80 μmol·L-1做了12个处理的试验,发现KT在4.61 μmol·L-1时水曲柳单芽基部分化率最低。所以,将水曲柳嫩茎尖诱导产生的单芽体切下接种到附加2-ip 15.80 μmol·L-1和KT 4.61 μmol·L-1的N6培养基上进行单芽试管保存的验证试验,重复3次,保存12个月统计的分化率仅为1.2%,继续保存至42个月后,测其平均分化率为0.5%以下,在估计区间内,且比所有试验Y值都低。
|
|

因此,水曲柳单芽体的最佳试管保存培养的适宜方案为: N6 + 2-ip15. 80 μmol·L-1 + KT 4.61 μmol·L-1。采用此培养基,在常温条件下,保存时间长达42个月以上,芽苗生长极其缓慢且形态、质量等均无明显变化(图 1c)。
解除保存后的单芽转接至N6 + 2-ip 9.84 μmol·L-1 + IAA 0.45 μmol·L-1培养基上进行单芽基部直接再生芽苗培养,培养25天后,单芽基部分化出大量粗壮的芽苗,且生长旺盛,形态和发育均未出现异常(图 1d),萌发率达95%以上。
3 结论与讨论本研究应用均匀设计法筛选出适宜水曲柳嫩茎尖离体培养和离体保存的激素组合方案,该法具有试验次数少、因素的水平可以适当调整等优点,大大缩短试验的摸索时间。
在水曲柳嫩茎尖直接再生芽苗的诱导培养基筛选试验中,当2-ip质量浓度低于8.36 μmol·L-1时嫩茎基部几乎不分化芽苗,超过11.86 μmol·L-1时芽苗诱导率下降,诱导率均在43.5%以下,说明过高的2-ip浓度对嫩茎基部直接再生芽苗起抑制作用。IAA质量浓度低于0.45μmol·L-1时嫩茎基部产生的芽苗生长极其缓慢,超过0.87 μmol·L-1时嫩芽基部产生大量愈伤组织; 生根培养基筛选试验结果表明,在1/4 N6培养基中附加较低浓度的IBA和NAA,加快生根速度,可能是低浓度的生长素促进根原基的形成。
植物种质保存研究对于生物多样性保护和新品种选育均具有十分重要的意义。目前,国内外已有很多关于植物种质保存方面的报道,大多数采取低温处理、玻璃化、包埋、低温结合弱光照等方法(钱剑林等,2004; Shawky et al., 2007; Acedo et al., 2005)。还有利用植物生长调节剂的不同作用机理,采取“矮化延缓生长”的方法对植物种质进行保存(艾鹏飞等,2004; 顾地周等,2008b; 顾地周等,2009b)。本研究对水曲柳试管苗的保存采取“激素超量”的方法,即在N6培养基中添加15. 80 μmol· L-12-ip并辅以4. 61 μmol·L-1的KT对水曲柳单芽体进行保存,这种方法可保存水曲柳单芽体达42个月以上,这可能是过量的激素抑制细胞的分裂,从而导致分化率下降等原因。通过对解除保存的水曲柳单芽进行再次诱导芽苗培养试验表明:解除保存后的单芽很快再次分化出大量粗壮的芽苗,形态和发育均未见异常。这说明采取“激素超量”法保存水曲柳试管苗是切实可行的。本试验成功建立水曲柳高效离体快繁和种质试管保存体系,为水曲柳这一珍贵树种资源的开发利用、工厂化育苗和种质资源保存奠定基础。
艾鹏飞, 罗正荣. 2004. 柿和君迁子试管苗缓慢生长法保存及其遗传稳定性研究[J]. 园艺学报, 31(4): 441-446. |
曹孜义, 刘国民. 2002. 实用植物组织培养技术教程[M]. 兰州: 甘肃科学技术出版社.
|
顾地周, 朱俊义, 姜云天, 等. 2008a. 东北刺人参组培快繁培养基的筛选[J]. 林业科学研究, 21(6): 867-870. |
顾地周, 孙忠林, 何晓燕, 等. 2008b. 牛皮杜鹃的组培快繁及种质试管保存技术[J]. 园艺学报, 35(4): 603-606. |
顾地周, 高捍东, 顾美影, 等. 2009a. 笃斯越桔离体培养及植株再生体系的建立[J]. 林业科学研究, 22(2): 226-229. |
顾地周, 罗微, 曹逊, 等. 2009b. 松毛翠的离体快繁体系建立及种质试管保存[J]. 林业科学, 45(7): 140-144. |
孔冬梅, 沈海龙, 冯丹丹, 等. 2006. 水曲柳体细胞胚与合子胚发生的细胞学研究[J]. 林业科学, 42(12): 130-133. DOI:10.3321/j.issn:1001-7488.2006.12.023 |
钱剑林, 朱旭东, 田松青, 等. 2004. 东方百合'Sorbonne'试管成球和低温处理的初步探讨[J]. 园艺学报, 31(6): 828. |
沈海龙, 赵霞, 邢朝斌, 等. 2005. 水曲柳扦插繁殖影响因子的分析[J]. 东北林业大学学报, 33(3): 5-6. |
谭燕双, 沈海龙. 2003. 水曲柳下胚轴的组织培养和植株再生[J]. 植物生理学通讯, 39(6): 623. |
汪松, 解焱. 2004. 中国物种红色名录.1卷[M]. 北京: 高等教育出版社: 304-464.
|
张惠君, 罗凤霞. 2003. 水曲柳未成熟胚的离体培养[J]. 林业科学, 39(3): 63-70. DOI:10.11707/j.1001-7488.20030310 |
张丽杰, 张丽玮, 冯丹丹, 等. 2007. 水曲柳腋芽离体快繁研究初报[J]. 植物研究, 27(3): 319-324. |
Acedo V Z, Arradoza C. 2005. In vitro conservation of yam germplasm[J]. Philippine Journal of Crop Science, 30(1): 112. |
Shawky B, Aly V I. 2007. In vitro conservation of globe artichoke(Cynara scolymus L.)germplasm[J]. International Journal ofAgriculture and Biology, 9(3): 404-407. |