 2010, Vol. 46
2010, Vol. 46文章信息
- 刘方炎, 李昆, 廖声熙, 崔永忠
- Liu Fangyan, Li Kun, Liao shengxi, Cui Yongzhong
- 濒危植物翠柏的个体生长动态及种群结构与种内竞争
- Interspecific Competition, Population Structure and Growth Dynamics of Endangered Calocedrus macrolepis
- 林业科学, 2010, 46(10): 23-28.
- Scientia Silvae Sinicae, 2010, 46(10): 23-28.
-
文章历史
- 收稿日期:2009-05-04
- 修回日期:2009-11-10
-
作者相关文章



2. 北京林业大学 北京 100083
2. Beijing Forestry University Beijing 100083
翠柏(Calocedrus macrolepis), 也称大鳞肖楠、长柄翠柏, 为柏科翠柏属常绿乔木, 属于国家2级珍稀濒危保护植物(傅立国, 1995)。翠柏属在全世界仅有2个种和1个变种, 间断分布于北美与中国。我国有1个种和1个变种, 即翠柏和台湾翠柏(Calocedrus macrolepis var.formosana)。翠柏主要分布在云南、贵州(三都)、广西(靖西)及海南岛等地, 常以纯林的形式存在。其木材优良, 纹理通直, 结构细密, 有光泽及香气, 耐腐, 是建筑、桥梁、家具的优良用材, 也是当地农村住房建筑材料中大柱、大梁的主要木材。从20世纪80年代起, 由于过量采伐以及生境的破坏, 翠柏资源数量出现了急剧减少。翠柏天然林几乎消失殆尽, 植株也逐渐稀少。主要存在状态是零星分布于房前屋后、田间地头、墓地以及道路两旁, 偶尔呈零散的小面积分布。滇中地区, 仅云南省易门县有较大面积的翠柏天然林, 而易门县也因此成立全国唯一的翠柏自然保护区(陈文红等, 2001)。虽然翠柏的濒危状况已引起社会各界的广泛关注, 但有关翠柏的研究报道甚少, 仅有部分学者针对其植株生长的环境(陈子牛, 1997), 植物区系、群落结构和物种组成(宁世江等, 1997; 陈文红等, 2001)等方面作了简要的介绍。有关其植株生长以及现有林分内的种内竞争状况等方面的资料尚未见报道。
植株的个体生长、种群结构与植株间的竞争强度存在着极为密切的关系, 一般认为, 植物间的竞争影响着植物的个体生长、形态结构以及存活状况, 也影响着其种群的空间分布格局、组成结构、动态变化及群落的物种多样性程度(Weiner, 1990; Masakado, 1997; Yokazaw et al., 1998)。而植株之间竞争的实质是对环境资源的争夺, 这种争夺必然会导致种群内部植株密度的改变, 即自疏现象的发生, 从而改变种群的数量结构(李博等, 1998)。在天然种群中, 竞争的最终结果是相邻植株达到相对稳定的最佳生态位(金则新等, 2004), 从而使种群数量维持在一个相对稳定的水平。研究天然林种群结构特征、植株个体生长状况以及种内竞争强度有利于探索翠柏植株数量减少以致濒危的原因, 从而指导人工抚育, 为翠柏的有效保护和利用提供有益的参考。
|
|
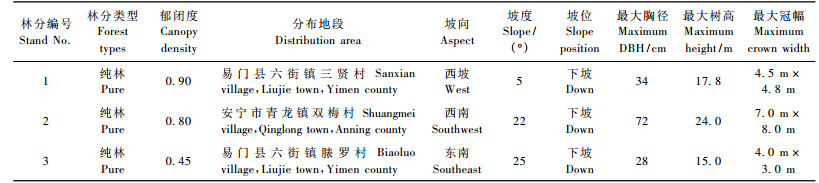
研究区位于滇中地区的安宁市青龙镇和易门县六街镇两地。两研究区均处于两市(县)交界处, 为中亚热带气候类型, 年平均气温15 ℃左右, 最冷月为1月, 月均温8 ℃, 最低气温-5 ℃; 最热月为6月, 月均温21.5 ℃, 最高气温34.6 ℃; 年平均降水量800 ~ 900 mm, 其中, 86%的降雨量集中在雨季(5—10月)。土壤为红壤, pH值4.5 ~ 5.5。
翠柏为中性偏阳性树种, 幼年耐荫, 以后逐渐喜光。耐旱性、耐瘠薄性均较强。常以小片纯林的形式存在, 乔木层中, 翠柏占95%以上, 为主要优势种。林中伴有少量杜英(Elaeocarpus sp.)、大头茶(Gordonia axillaris)、粗叶水锦树(Wendlandia scabra)、云南油杉(Ketelleeria evelyniana), 云南松(Pinus yunnanensis)、白皮柯(Lithocarpus dealbatus)、光叶石栎(L.mairei)等乔木树种。林下植物有云南含笑(Michelia yunnanensis)、薄叶鼠李(Rhamnus leptophylla)、小铁仔(Myrsine africana)、小叶女贞(Ligustrum quihoui)、细柄十大功劳(Mahonea gracilipes)、香茶菜(Rabdosia amethystoides)、火绒草(Leontopodium leontopodioides)、香青(Anaphalis sp.)、云南兔耳风(Ainsliaea yunnanensis)、千里光(Senecio scandens)、茅叶荩草(Arthraxon lanceolatus)、拳参(Polygonum bistorta)、委陵菜(Potentilla chinensis)等。翠柏更新良好, 母树周围幼苗最多。大部分幼苗随着幼树喜光阶段的来临, 耐荫性的减退而逐渐死亡。
1.2 野外调查 1.2.1 样地设定本研究在对翠柏资源全面踏查的基础之上, 于2007年11月分别在滇中地区的安宁市和易门县两地, 选取翠柏植株保存相对完好, 面积在2 000 m2以上不同郁闭程度的林分3块。每一林分中设样地3个, 样地面积为20 m × 30 m, 共设样地面积5 400 m2。对样方内所有高度在1.5 m以上植株进行每木检尺, 记录其胸径、树高和冠幅。每一样地内均设5个1 m × 1 m的样方, 调查1.5 m以下幼小植株及幼苗数量。高大乔木树高采用威特OPTI-LOGIC(600XV)激光测高仪测得; 林分郁闭度的测定采用测线法, 即在每一林分内垂直等高线量取40 m测线, 观察树冠投影在测线上的长度, 计算各树冠投影长度之和与总长度之比(即林分郁闭度), 重复3次求平均值。
1.2.2 数据分析1) 胸径与树高关系 对所调查的497棵翠柏植株, 以胸径小于2 cm的植株为1个级别, 然后每隔1 cm划分为1个级别(径级不存在的即空出)。求出每一径级中胸径和树高的平均值, 利用胸径平均值计算出胸高断面积, 找出胸高断面积与对应树高平均值之间关系, 然后利用相应软件进行拟合。
2) 径级结构 在径级结构划分时, 将胸径小于2 cm的植株作为1个径级, 即径级为 < 2 cm。胸径大于等于2 cm的植株每隔4 cm划为1个径级, 即胸径为2 cm≤D < 6 cm的植株径级为4 cm, 6 cm≤ D < 10 cm的植株径级为8 cm, 依此类推。按此标准统计植株个体数, 然后以径级为横坐标, 植株个体数为纵坐标, 作出各样地翠柏径级结构图。
3) 种内竞争强度 在众多林木竞争指数的模型中, Hegyi提出的有关距离的预测模型得到了较多的认可和广泛的应用(张思玉等, 2001a; 金则新等, 2004), 同时, 该模型在野外调查过程中相对便捷。本文即采用Hegyi提出的竞争指数计算单木及各样地林分个体之间竞争状况。根据林分内胸径最大翠柏植株的冠幅大小和遮荫程度确定以对象木为中心, 半径为8 m样圆内所有翠柏为竞争木, 测定对象木及竞争木胸径以及竞争木距相应对象木距离。其具体计算方法如下:

|
式中, CI为竞争指数, 其值越大, 对象木受到竞争木的竞争越剧烈; Di为第i棵对象木的胸径; Dj为第j棵竞争木的胸径; Lij为第i棵对象木与第j棵竞争木之间的距离; N为竞争木的株数。
单木竞争指数的调查与计算方法为:在所有样地中随机选择胸径 < 10, 11 ~ 20, 21 ~ 30, 31 ~ 40, 41 ~ 50, 51 ~ 60以及> 61 cm的植株各5株作为对象木, 调查并计算各自竞争指数, 求出各径级单株平均竞争指数。不同林分之间种内竞争指数调查与计算方法为:在每一林分3个样地中各随机选择胸径 < 10, 11 ~ 20, 21 ~ 30, 31 ~ 40, 41 ~ 50, 51 ~ 60以及> 61 cm的植株各3株作为对象木(径级不存在的不计算), 调查并计算竞争指数, 求出各样地单株平均竞争指数, 然后求出各林分的单株平均竞争指数。
数据处理采用SPSS 11.5软件中的单因素方差分析One-way ANOVA, 作图及数据拟合采用SIGMAPLOT 9.0软件完成。
2 结果 2.1 植株个体生长动态天然林中, 翠柏幼苗生长初期, 高生长相对胸高断面积的增长来说, 优势十分明显。胸径在0.5 ~ 1.0 cm的植株, 其高度通常可达1.5 ~ 2.5 m。通过对翠柏天然林中植株(胸径在0.5 ~ 72 cm)胸高断面积与树高关系的研究发现, 胸高断面积与树高之间存在着一种显著的指数函数关系(图 1)。回归方程的相关系数R2 = 0.981, 具有较高的可信度。从图中可以看出, 翠柏植株在胸高断面积不断增长的过程中, 其高生长也在不断持续。在胸高断面积增长的初期, 其高生长增加显著, 但当胸高断面积约0.102 m2(胸径约36 cm)、树高约19 m之后, 其胸高断面积开始迅速增长, 而高生长增长趋于平缓。表明翠柏植株在树高达到约19 m之前主要进行的是高生长, 之后, 其胸径生长占有较大优势, 树高生长开始慢慢减弱直到停止。在翠柏资源全面踏查过程中也发现, 翠柏植株树高超过20 m的较少。在楚雄彝族自治州禄丰县土官镇烈士陵园中生长的翠柏千年古树其胸径近200 cm, 而树高也仅约24 m。

|
图 1 翠柏植株胸径与树高关系 Figure 1 Relationships between tree height and DBH of C.macrolepis |
种群年龄结构的分析是探讨种群动态特征的一种有效的方法。而对稀有濒危植物而言, 利用径级结构代替其年龄结构分析树种的结构和动态变化特征是一种非常实用而可行的方法(张文辉等, 2004; 段仁燕等, 2007); 同时, 径级结构也是反映种群结构稳定性的一个重要指标。
从图 2可以看出, 滇中高原翠柏天然林植株径级分布极不规律, 均呈现出连续状, 除林分1以外, 林分2, 3均没有表现出一个完整的更新系列。表明现存的少量翠柏天然林长期以来也均受到不同程度的各种干扰因素的影响, 林分结构不完整, 种群生长环境出现了较大程度的破碎化, 种群稳定性不高。其中, 林分2中径级12 cm(胸径为10 ~ 14 cm)以及林分3中径级8, 12 cm(胸径为6 ~ 14 cm)的植株个体数均为零, 出现了明显的断裂, 表明这2个林分内的翠柏曾受到极为严重外部干扰, 部分胸径大小相对一致的植株个体大量消失, 或者其幼苗更新曾经出现了阻断, 种群生存压力较大。但从各林分中不同径级植株所占的比例来看, 3个林分中径级为8 cm以下植株所占比例均较大, 分别达到了39.3%, 48.9%和84.5%。表明近期以来, 3个林分中翠柏林下幼树更新能力均相对良好。而且, 在3个不同郁闭度的林分中, 尤其是林分3中, 胸径小于2 cm的幼小植株数均多于其他径级植株, 表明现有翠柏天然种群中均不缺乏更新资源, 翠柏天然种群自然更新能力均较强, 林下幼苗天然更新状况均良好, 如不出现较大程度的外来干扰, 现有翠柏天然种群能够出现较大程度的增长。

|
图 2 3个林分植株径级结构特征比较 Figure 2 A comparison of plant diameter in three stands |
植物物种之间的竞争能力主要取决于其生态习性、生态幅度以及其在生态系统中的生态位, 生态习性越接近, 植株之间的竞争越激烈。对于纯林中的植株个体来说, 植株之间的生态习性完全一致, 其种群中植株的平均竞争强度主要取决于植株的密度和个体大小。从各样地翠柏植株单株平均竞争强度比较可以看出(图 3), 3个林分中翠柏纯林的种内竞争强度存在着显著差异。其中, 林分1中翠柏植株的平均竞争强度最大, 其次是林分2, 竞争强度最小的是样地3。数值变化在1.97 ~ 4.31。表明现有零星分布的小片翠柏纯林受干扰的程度以及林地存在的时间不同, 以致种群平均密度和植株的大小存在较大差异。

|
图 3 各样地内植株平均单株竞争指数比较 Figure 3 Comparison of average competition indexes of C.macrolepis in different sample plots 不同字母表示竞争指数存在显著差异Different letters indicate significant different at the level of 0.05。下同The same below. |
对于单株植物来说, 对象木所受竞争强度与其胸径的大小存在一定关系(张池等, 2006)。从翠柏不同胸径植株(大于5 cm)的竞争指数曲线图中可以看出(图 4), 翠柏植株受到的竞争强度随着其胸径的增大而逐渐减小。其中, 胸径在 < 10 cm至31 ~ 40 cm的植株, 竞争指数随胸径增大呈现急剧下降的趋势, 但胸径在31 ~ 40 cm以后, 竞争指数下降趋于平缓。同时, 从不同胸径植株间竞争指数的方差分析来看, 胸径为 < 10, 11 ~ 20, 21 ~ 30以及31 ~ 40 cm的植株竞争指数之间存在着显著差异, 而31 ~ 40, 41 ~ 50和51 ~ 60 cm之间差异不显著。表明翠柏植株胸径在31 ~ 40 cm时, 其竞争指数出现较为明显的变化, 胸径在31 ~ 40 cm以下的植株受到其他植株竞争的作用较强, 不同胸径间植株的竞争作用差异较大, 但31 ~ 40 cm以上的植株受到其他植株的竞争压力相对较弱, 且差异不大。

|
图 4 不同胸径翠柏植株竞争强度比较 Figure 4 Comparison of competition indexes of individual trees with different DBH |
天然林中, 幼苗和幼树的生长对森林生态系统的稳定性具有非常重要的作用。其中, 主要优势种植物种子的产生、萌发以及幼苗的定居和生长状况均能够影响到种群结构动态及其持续稳定的发展潜力。因此, 维持种群稳定性的关键是要为种子萌发、幼苗定居和生长创造适宜条件。其中任何一个环节出现中断, 适宜条件遭到破坏, 整个种群生态系统稳定性将会被打破, 从而会导致种群的濒危, 甚至灭绝。现有许多濒危植物, 如太白红杉(Larix chinensis)、银杉(Cathaya argyrophylla)以及秦岭冷杉(Abies chensiensis)等物种的致危原因即是天然条件下种群结实量少、种子生活力差等因素致其种子转换成幼苗的概率过低所致(张文辉等, 2004; 2006;谢宗强等, 1999)。但从翠柏天然林中幼苗更新现状来看, 翠柏的致危因子与以上可能有着较大的差别, 其林下种子自然萌发更新能力极强。天然林中也能发现林下幼苗十分密集(宁世江等, 1997), 单位面积(1 m2)1年生幼苗株数可达100株以上, 平均株高可达5 cm(陈子牛, 1997)。这与其种子产量大、出苗率高以及幼苗阶段具有较好的耐荫性有着较为密切的关系。同时, 从现存林分中翠柏植株的径级结构也能看出, 幼小植株在3个林分中所占比例均较大, 翠柏天然林种群数量均呈现出增长的趋势。但随着幼小植株径级的进一步增大, 数量均出现明显减小(图 2)。表明林下翠柏幼树在生长过程中均受到某一限制因子的制约, 出现植株大量死亡现象。从翠柏植株的个体生长过程来看, 其生长初期主要进行着高生长, 其目的是以最快的速度争取到林分上层充足的光照。从3个林分中幼树数量的比较中也可发现, 林分郁闭度越高, 树高在1.5 m以下植株和胸径小于5 cm植株数量越少, 反之则越多。由此可知, 林下光照强弱可能是影响其幼苗更新极为重要的因子之一。而林分1, 2中的植株密度或郁闭度可能过高, 影响林下幼树的进一步生长。
通常, 林下幼苗更新与森林光环境异质性特别是光照强度间具有较强的关联性(Scholes et al., 1997; Thomas et al., 1998), 不同树种幼苗不仅对光的需求不同, 对光环境变化的反应以及适应能力也不一样(Swaine et al., 1988)。翠柏幼苗虽具有较好的耐荫性, 但随着植株的长大, 其耐荫性逐渐减弱, 喜光性逐渐增强(陈子牛, 1997)。由于翠柏植株的树冠常常较为浓密, 林下郁闭度极高, 自然生长状况下常在0.90以上。其产生的影响是使其他阔叶树种幼苗无法生长, 维持了种群的生存空间, 但也导致自身幼苗生长到一定程度以后, 林下光照无法满足其生长需要, 大部分幼苗出现死亡的现象。同时, 由于翠柏幼苗具有较强的耐荫性, 其在光照强烈的开阔地, 如林缘等环境中生存困难, 从而较难拓展其生长领地。因此, 导致翠柏濒危的主要因子可能不是其种子产量低、生活力差等自身繁殖更新能力的退化, 而是其自然分布区的立地环境遭到破坏以后, 残存翠柏植株产生的种子在林缘处萌发以后, 其幼苗在强烈光照下容易死亡造成的。因此, 在翠柏的保护过程中, 应加强现有翠柏林缘生境的保护, 从林缘开始逐渐拓展其生存空间, 才能使其脱离濒危状态。
3.2 种内竞争与种群稳定性植株间的竞争是一种普遍的现象。竞争的结果不仅影响个体的生存、生长和繁殖, 而且影响种群空间分布以及动态变化等(Weiner, 1990; Yokazawa et al., 1998)。与混交林内植株相比较, 纯林内植株之间由于具有相同的生态习性和生态幅度以及共同的可利用资源, 从而比任何一种具有伴生树种林分的竞争都要剧烈。自1961年Harper提出邻体干扰问题以来, 许多植物生态学家推出描述林木间竞争强度的数量指标, 即竞争指数。竞争指数反映的是树木个体生长与生存空间的关系, 同时也反映树木对环境资源的需求与现实情况下对环境资源的占有量之间的关系(张思玉等, 2001b)。在众多的竞争指数中, Hegyi的竞争指数模型被认为是最能反映种群竞争现状的模型(邹春静等, 1998)。它主要考虑2个因素, 即植株的胸径大小比值以及植株之间的距离, 也就是植株大小和生长密度2因子。植株个体增大、密度增高以后, 个体对资源的占有量急剧减少, 相互之间的竞争作用将增强。林分1中成年植株密度最大, 种群内植株平均竞争指数也最大。但从翠柏植株不同胸径个体竞争能力来看, 该样地中大量植株正处于激烈竞争阶段, 最大植株胸径也没有达到竞争趋于平缓的31 ~ 40 cm, 因此, 该样地翠柏种群的竞争仍会加强。这种竞争逐渐加强的趋势对密度较高的翠柏纯林来说可能是一种必然过程。通常来说, 当密度较小、竞争指数较小时, 个体死亡概率很小, 且随着竞争指数的增大变化不大; 当竞争指数超过一定数值时, 多数天然林树种, 如甜槠(Castanopsis eyrei)、日本落叶松(Larix kaempferi)等植株的死亡概率会急剧增大, 从而会出现一个“自疏”过程(金则新等, 2004; 张泽浦等, 2000)。从径级结构和林龄组成来看, 林分2的翠柏种群应处于这个过程的末期。而林分3是人为因素影响严重但并没有出现毁灭性的破坏后幼苗幼树迅速生长的情况。后两者共同特点是林地破碎化程度较高, 林内成年植株密度较低, 林隙大量存在, 郁闭度相对较低, 但幼苗幼树所占比例较高。从3个林分植株生存状况和特征来看, 在没有外来干扰的情况下, 现有翠柏种群可以完成自然更新过程, 从而使种群达到相对的稳定。从不同胸径翠柏竞争指数的变化来看, 平均胸径为37 cm左右可能也是这一“自疏”过程拐点。因此, 在经营翠柏人工林时, 可以参照其天然林生长特征, 进行合理的经营和管理。
陈文红, 税玉民, 王文, 等. 2001. 云南易门翠柏和黄杉的群落调查及保护[J]. 云南植物研究, 23(2): 189-200. |
陈子牛. 1997. 滇中翠柏纯林的生态研究[J]. 昆明师范高等专科学校学报:自然科学版, 12(2): 15-22. |
段仁燕, 王孝安, 黄敏毅, 等. 2007. 太白红杉混交林径级结构与竞争的关系[J]. 生态学报, 27(11): 4919-4924. DOI:10.3321/j.issn:1000-0933.2007.11.064 |
傅立国. 1995. 中国植物红皮书(第1册)[M]. 北京: 科学出版社.
|
金则新, 朱小燕, 林恒琴. 2004. 浙江天台山甜槠种内与种间竞争研究[J]. 生态学杂志, 23(2): 22-25. |
李博, 陈家宽, 沃金森A R. 1998. 植物竞争的研究进展[J]. 植物学通报, 15(4): 18-29. |
宁世江, 赵天林, 唐润琴, 等. 1997. 木论喀斯特林区翠柏群落学特征的初步研究[J]. 广西植物, 17(4): 321-330. |
谢宗强, 陈伟烈. 1999. 中国特有植物银杉的濒危原因及保护对策[J]. 植物生态学报, 23(1): 1-7. |
张思玉, 郑世群. 2001a. 笔架山常绿阔叶林优势种群内种间竞争的数量研究[J]. 林业科学, 37(专1): 185-188. |
张思玉, 郑世群. 2001b. 永定桫椤群落的结构特征[J]. 植物资源与环境学报, 10(3): 30-44. |
张文辉, 王延平, 康永祥. 2004. 濒危植物太白红杉种群年龄结构及其时间序列预测分析[J]. 生物多样性, 12(3): 361-369. |
张文辉, 许晓波, 周建云. 2006. 濒危植物秦岭冷杉生殖生态学特征[J]. 生态学报, 26(8): 2417-2424. |
张泽浦, 方精云, 菅诚. 2000. 邻体竞争对植物个体生长速率和死亡速率概率的影响:基于日本落叶松种群试验的研究[J]. 植物生态学报, 24(3): 340-345. |
邹春静, 徐文铎. 1998. 沙地云杉种内、种间竞争的研究[J]. 植物生态学报, 22(3): 269-274. |
Masakado K. 1997. Exploitative competition and ecological effective abundance[J]. Ecologiacal Modelling, 94(2/3): 125-137. |
Scholes J D, Press M C, Zipperlen S W. 1997. Differences in light energy utilisation and dissipation between dipterocarp rain forest tree seedlings[J]. Oecologia, 109(1): 41-48. DOI:10.1007/s004420050056 |
Swaine M D, Whitmore T C. 1988. On the definition of ecological species groups in tropical rain forests[J]. Vegetatio, 75(1/2): 81-86. |
Thomas T L, Martin J L. 1998. Diverse responses of maple saplings to forest light regimes[J]. Annals Botany, 82(1): 9-19. DOI:10.1006/anbo.1998.0644 |
Weiner J. 1990. A symmetric competition in plant population[J]. Trends in Ecology and Evolution, 5(11): 360-364. DOI:10.1016/0169-5347(90)90095-U |
Yokazawa M, Kubota Y, Hara T. 1998. Effects of competition mode on spatial pattern dynamics in plant communities[J]. Ecological Modelling, 106(1): 1-16. DOI:10.1016/S0304-3800(97)00181-6 |