 2010, Vol. 46
2010, Vol. 46文章信息
- 杨小林, 赵垦田, 马和平, 禄树晖, 罗健
- Yang Xiaolin, Zhao Kentian, Ma Heping, Lu Shuhui, Luo Jian
- 拉萨半干旱河谷地带的植被数量生态研究
- Ecological Studies on Vegetation Quantity in the Semi-Arid Valley Region of Lasa
- 林业科学, 2010, 46(10): 15-22.
- Scientia Silvae Sinicae, 2010, 46(10): 15-22.
-
文章历史
- 收稿日期:2009-09-11
- 修回日期:2010-06-14
-
作者相关文章



当今, 由于人类活动的影响, 气候变化、生物多样性丧失、资源枯竭和生态环境退化, 导致了一系列的环境失调问题, 并严重威胁到人类的生存和社会的可持续发展, 在全球气候变暖的背景下, 河谷地带的干旱化过程有进一步加剧的趋势(陈灵芝, 1993; 杨继平, 2000; Lomolino, 2001), 河谷地带人口、资源、环境与发展之间的矛盾日趋突出。人类活动对环境的影响以及退化生态系统的恢复和重建成为现代生态学研究最引人注目的领域之一(马世骏, 1990; 章家恩等, 1997)。作为研究退化生态系统恢复与重建问题的恢复生态学受到世界各国的普遍重视, 并成为现代生态学研究的主要领域(章家恩等, 1997; 1999; 周国逸, 1998)。
植物群落的分类是植物群落学中最复杂的问题之一。植被数量分类和排序研究, 多从生态学的群落概念出发, 以群落为研究单位进行类型的划分, 即乔、灌、草各层物种均统计在内, 它的优点在于可以进行群落类型划分及环境解释分析(江洪, 1994; 张金屯, 1995; 张峰等, 2000)。目前, 在我国曾对许多地区的植被进行了数量分类(祝廷成等, 1988; 席跃翔等, 2004; 胡理乐等, 2005; 郭东罡等, 2005; 代力民等, 2008), 因此, 学者们将数量分析方法引入传统的植物群落分类(Zdravko, 2005; 熊咏梅等, 2008)。
拉萨河谷是西藏“一江两河”的重点区域之一, 其生态环境的退化影响着西藏高原国家生态安全屏障的构建。本文应用TWINSPAN和DCA多元分析的方法, 以乔、灌、草各层物种的生态重要值为指标, 对半干旱河谷区域内样方进行数量分类和排序研究, 旨在深入理解高原半干旱河谷区域的植被空间分布及植被的生态适应特性, 为高原河谷地带植被的恢复和重建提供理论依据。
1 研究区自然概况拉萨半干旱河谷地处雅鲁藏布江支流拉萨河流域, 喜马拉雅山北侧, 受下沉气流的影响, 全年多晴朗天气, 冬无严寒, 夏无酷暑, 属高原季风半干旱气候。区内气候温暖、干燥, 年均气温7.4 ℃, 年平均相对湿度30% ~ 50%, 降水量200 ~ 500 mm, 干燥度1.5 ~ 10, 干湿指数3 ~ 7, ≥10 ℃积温2 177 ℃, 无霜期133天, 全年日照时数3 000 h以上(青藏高原综合科学考察队, 1985)。本研究区位于拉萨河中下游河谷地带, 29° 15′—30° 00′ N, 90° 45′—91° 50′ E, 面积330.4 km2, 为拉萨半干旱河谷的典型地带, 海拔3 650 ~ 5 000 m, 河谷较宽, 发育有河漫滩、堆积阶地、洪积扇、河谷盆地和山地等地貌类型。属于雅鲁藏布江中游谷地亚高山灌丛、草原亚区植被区(青藏高原综合科学考察队, 1988)。
拉萨半干旱河谷植被类型主要为亚高山灌丛和草甸植被以及河谷人工林群落。在河谷河漫滩天然分布有沙棘(Hippophae rhamnoides)、小花水柏枝(Myricaria wardii)、大红柳(Salix cheilophila var. microstachyoides)群落, 以及以杨树(Populus spp.)、柳树(Salix spp.)为主的人工林群落。在海拔4 000 m以下, 主要分布以砂生槐(Sophora moorcroftiana)、架棚(Ceratostigma minus)等组成的落叶灌丛; 海拔4 000 ~ 4 300(4 500)m, 植被有绢毛蔷薇(Rosa sericea)、高山绣线菊(Spiraea alpina)、小檗(Berberis sp.)等亚高山灌丛和藏沙蒿(Artemisia wellbyi)、丝颖针茅(Stipa capillacea)、固沙草(Orinus thoroldii)等组成的小半灌木草原群落。海拔4 300(4 500) ~ 4 800 m, 香柏(Sabina pingii var.wilsonmii)灌丛(阳坡)和杜鹃(Rhododendron spp.)、三叶金露梅(Potentilla fruticosa var.tangutica)、鬼箭锦鸡儿(Caragana jubata)等组成的常绿针叶和常绿革叶灌丛等; 局部石砾质的地段还分布有小片垫状植被。海拔4 800 m以上主要以高山嵩草(Kobresia pygmaea)为主组成的高山草甸植被, 阴坡和阳坡植被类型基本相近, 并有片段的垫状植物群落。5 000 m以上为高山流石滩稀疏植被(高山冰缘植被)。
2 研究方法 2.1 研究区的确定本研究以拉萨河流域半干旱河谷地区的自然植被分布为依据, 选择河谷最低海拔3 650 ~ 5 000 m, 海拔垂直带谱1 350 m的范围, 包括拉萨市北山(阳坡)、南山(阴坡)和河谷地带进行调查研究。
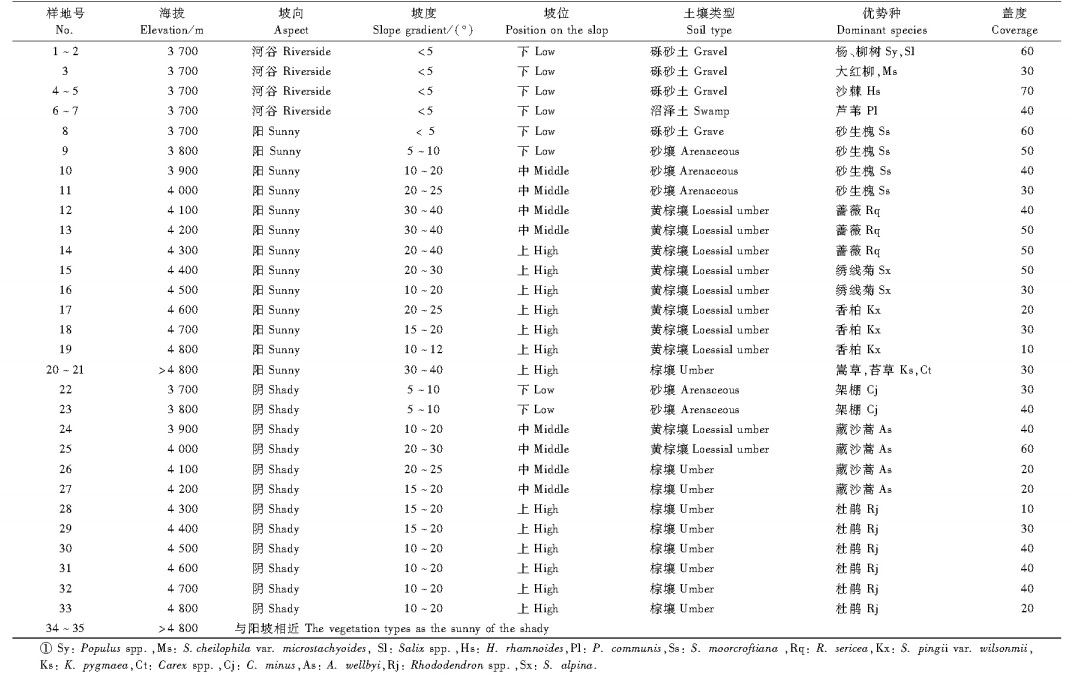
2.2 外业调查分别在阳坡(北山)和阴坡(南山)按垂直高差100 m设置典型样地, 同时在河谷地带按照植被类型设置典型样地, 阳坡与阴坡设置样地28块, 河谷地带设置7块, 共获得典型样地35块。根据植被类型设置样地大小, 拉萨河谷地带植被主要以灌丛和灌丛草原为主, 样地大小设置为10 m × 10 m, 在每个样地内沿对角线设置5个草本样方, 共获得草本样方175块。在每个样地和样方内, 分别记录植物的种类、株数、高度、盖度等, 同时记录样地的海拔、坡向、坡度和土壤等环境指标。各样地概况见表 1所示。
|
|
本研究应用调查样方中的乔木、灌木和草本种统计数据, 按照重要值的计算方法:乔木层:Ⅳ = (相对高度+相对显著度+相对频度)/3;灌木层: Ⅳ =(相对高度+相对盖度+相对频度)/3;草本层; Ⅳ =(相对高度+相对盖度+相对频度)/3。
重要值取值范围为0 ~ 1。在研究区域不同海拔梯度上, 共取得35块样地, 记录了196种子植物, 形成35 × 196维的数据矩阵, 按样方和物种分别进行数量分类和排序分析。样方分类采用物种重要值数据, 物种生态种组划分采用二元属性数据, 即(0, 1)数据。分类采用VESPAN软件包中的二元指示种分析TWINSPAN(two-way indicators species analysis)(Hill, 1997), 排序采用CANOCO软件包标准程序的除趋势对应分析DCA(detrended correspondence analysis)(ter Braak, 1988)。
2.4 植被分类单位和命名植被分类单位和命名依据青藏高原综合科学考察队(1988)的分类系统。其中采用植被型、群系、群丛为主要的植被分类单位。
对群系和群丛的命名采用优势种命名法, 用群落各层优势种的拉丁学名来命名, 依优势度的大小依次列出主要的优势种, 并在学名之后加上分类单位名称的全称或缩写。
3 结果与分析 3.1 植被数量分类结果图 1为拉萨半干旱河谷研究区域所有植物重要值的35个样地TWINSPAN分析结果, 将拉萨半干旱河谷地带的主要植被划分成4个植被型组, 即阔叶林、灌丛、草甸和沼泽; 7个植被型; 14个群系; 根据分类结果结合调查将14个群系划分成16个群丛。具体结果见表 2。

|
图 1 拉萨半干旱河谷地带的35个样地TWINSPAN分析结果 Figure 1 TWINSPAN classification results of the 35 plots of semi-arid valley region of Lasa |
|
|

1) 杨树+劲直黄芪群丛, 样地号1。拉萨河谷地带人工林群落, 纯林, 结构简单, 栽植密度大, 郁闭度一般在0.7以上。栽培杨树主要有银白杨(Populus alba)、北京杨(P.pekinensis)、藏川杨(P. szccuanica var.tibetica)等; 林下植物种类单一, 主要为劲直黄芪(Astragalus strictus)、钉柱委陵菜(Potentilla saundersiana)、猪毛蒿(Artemisia scoparia)等, 盖度10% ~ 15%。
2) 柳树+玫红野青茅+长尖莎草群丛, 样地号2。拉萨河谷地带人工林群落, 海拔3 700 m左右, 纯林, 郁闭度0.5左右; 草本层较杨树林丰富, 主要为玫红野青茅(Deyeuxia rosea)、长尖莎草(Cyperus cuspidatus)、柔毛委陵菜(Potentilla saundersiana)、垂穗披碱草(Elymus nutans)、小球花蒿(Artemisia moorcroftiana)等, 在水位较高的地带, 草本盖度90%以上, 可以提供饲草。栽培种有垂柳(S. babylonica)、长蕊柳(S.longistaminea)等。
3) 沙棘+白草群丛, 样地号4, 5。拉萨河谷天然林群落, 海拔3 700 ~ 3 800 m拉萨河流域的河漫滩, 以沙棘为单优势种, 平均高度4 ~ 5 m, 平均胸径10 ~ 15 cm, 盖度40% ~ 70%;草本层主要以白草(Pennisetum flaccidum)、蕨麻委陵菜(Potentilla anserina)、香藜(Chenopodium botrys)、苔草(Carex spp.)、大籽蒿(Artemisia sieversiana)等, 草本层盖度80%, 以及铁线莲(Clematis sp.)。
4) 大红柳+小花水柏枝群丛, 样地号3。天然块状分布拉萨河谷中游的河漫滩, 呈灌木状, 群落平均高度1.5 m, 盖度30% ~ 40%, 草本层主要以早熟禾(Poa annus)、香藜、白草、拉萨黄芪(Astragalus lasaensis)等, 草本层盖度20%。
5) 砂生槐+架棚+藏沙蒿群丛, 样地号8, 9, 10, 11。分布海拔3 700 ~ 4 000 m山麓及阳坡(北山), 坡度5 ~ 25°, 灌丛盖度30% ~ 60%, 以砂生槐为主, 平均高度60 cm, 重要值0.56, 还有架棚、藏沙蒿、醉鱼草(Buddleja sp.)等; 草本层主要有白草、固沙草、丝颖针茅、早熟禾、须芒草(Andropogon yunnanensis)、喜马拉雅草沙蚕(Tripogon hookerianus)、香藜、圆齿红景天(Rhodiola crenulata)等, 草本盖度40%。
6) 绢毛蔷薇+小叶栒子+醉鱼草群丛, 样地号12, 13, 14。分布海拔4 000 ~ 4 300 m阳坡, 坡度20 ~ 40°, 灌丛盖度40% ~ 50%, 以绢毛蔷薇为主, 平均盖度160 cm, 重要值0.51, 其次为山岭麻黄(Ephedra gerardiana)、小檗混生有藏沙蒿、醉鱼草等灌木; 草本层主要为高山蒿草、翼首花(Pterocephalus hookeri)、冰岛蓼(Koenigia islandica)、扁芒草(Danthonta schneideri)、固沙草、须芒草(Antropogon gayanus)、拉萨黄芪、白草、直立点地梅(Androsace erecta)、青藏狗哇花(Heieropappus boweri)、毛果草(Lasicaryum)等, 草本盖度50%。
7) 高山绣线菊+山岭麻黄+小檗群丛, 样地号15, 16。分布海拔4 300 ~ 4 500 m阳坡, 坡度10 ~ 30°, 灌丛盖度30% ~ 50%, 以高山绣线菊为主, 伴生有山岭麻黄、小檗等灌丛; 草本主要由拉萨黄芪、青藏狗哇花(Heteropappus sp.)、尼泊尔大丁草(Leibnitzia nepalensis)、大花嵩草(Kobrasia macrantha)、高山嵩草、猪毛蒿(Artemisia scoparia)、冰原白蒿(Artemisia stracheyi)、鸢尾(Iris decora)、冰岛蓼(Koenigia sp.)、龙胆(Gentiana spp.)、蓝钟花(Cyananthus hookeri)、垂穗披碱草(Elymus nutans)等组成, 盖度50% ~ 60%。
8) 大果香柏-小檗+大花嵩草群丛, 样地号17, 18, 19。分布海拔4 500 ~ 4 800 m阳坡, 坡度10 ~ 25°, 盖度20% ~ 30%, 以大果圆柏和小檗为主, 高度100 ~ 200 cm, 混生有香柏(Sabina pingii var. wilsonmii)、金露梅(Potentilla fruticosa var.arbuscula)等; 草本层以大花嵩草为主, 伴生有早熟禾、羊茅(Festuca ovina)、木根香青(Anaphalis xylorhiza)、拉萨黄芪、螃蟹甲(Phlomis younghusbandii)、唐松草(Thalictrum ruiaefolium)等, 草本盖度30% ~ 40%。
9) 架棚+小叶丁香群丛, 样地号22, 23。分布海拔3 700 ~ 3 900 m阴坡(南山), 坡度5 ~ 10°, 盖度40%, 以架棚为主, 高度30 ~ 50 cm, 伴生有小叶丁香(Syringa microphylla); 草本层由丝颖针茅、扁芒草(Danthonia schneideri)、黄芪、丛茎滇紫草(Onosma waddellii)、卷柏(Selaginella pulvinata)、藏沙蒿、直立点地梅(Androsace erecta)、固沙草、马先蒿(Pedicularis spp.)等组成, 盖度40% ~ 60%。
10) 藏沙蒿+川藏香茶菜+小叶香茶群丛, 样地号24, 25, 26, 27。分布海拔3 900 ~ 4 300 m阴坡, 坡度10 ~ 25°, 盖度50%, 以藏沙蒿为主, 高度40 ~ 80 cm, 伴生有川藏香茶菜(Rabdosia pseudoirrorata)、小叶香茶菜(Rabdosia parvifolia)、固沙草、肋柱花(Lomatogonium thomsonii)、翼首花、卷柏(Selaginella sp.)、冰岛蓼、丝颖针茅、冻源白蒿等。
11) 杜鹃+金露梅+鬼箭锦鸡儿群丛, 样地号28, 29, 30, 31, 32, 33。分布海拔4 300 ~ 4 800 m阴坡, 以微毛樱草杜鹃(R.Primulaeflorum var. cephalanthoidea)占优势, 伴生有三叶金露梅、鬼箭锦鸡儿(Caragana jubata)以及少量的蒙古绣线菊(Spiraea mongolica), 灌木丛高60 ~ 70 cm, 盖度10% ~ 40%;草本层主要有嵩草(Kobresia spp.)、苔草(Carex spp.)、羊茅(Festuca sp.)、高山唐松草(Thalictrum alpinum)、虎耳草(Saxifraga sp.)、垫状点地梅(Androsace spp.)等, 草本层盖度50%。
12) 高山嵩草+大花嵩草+点地梅群丛, 样地号20, 34。海拔4 800 ~ 5 000 m阴、阳坡, 在海拔4 800 m以上阴坡和阳坡植被群落相近, 以高山嵩草为主要成分, 伴生有大花嵩草(Kobresia macrantha)、点地梅, 常见的种类有火绒草(Leonlopodium sp.)、木根香青(Anaphalis xylorhiza)、委陵菜(Potentilla spp.)、山莓草(Sibbaldia sp.)、蓼(polygonum sp.)、冰川棘豆(Oxytropis glacialis)、藏玄参(Oreoslen wattii)、紫花点地梅(Androsace selago)、蚤缀(Arenaria spp.)等, 盖度60%。
13) 苔草+红景天+垂头菊群丛, 样地号21。海拔5 000 m以上, 阴、阳坡, 属高山冰缘(Cremanthodium sp.)植被, 在流石扇缘地常见由苔草、红景天(Rhodiola spp.)、垂头菊(Cremanthodium sp.)组成的聚群, 伴生有马先蒿(Pedicularis sp.)、报春(Primula spp.)、龙胆(Gentiana sp.)、高山嵩草、虎耳草等, 盖度40%。
14) 圆穗蓼+凤毛菊群丛, 样地号35。海拔5 000 m以上, 阴、阳坡, 属高山冰缘植被, 在流石滩背风、潮湿的小环境, 通常由圆穗蓼(Polygonum macrophllum)、凤毛菊(Saussurea sugoo)为主要成分的聚群, 常见的种类有珠芽蓼(P.viviparum)、雪莲(Saussurea spp.)、红景天、虎耳草、报春等矮小草本植物, 盖度20% ~ 30%。
15) 芦苇+香蒲+藨草群丛, 样地号6。为拉萨河谷主要的湿地植被群落, 分布海拔3 700 m左右, 植物以高原特有的水生及半水生和草甸植物组成, 总盖度在90%左右, 群落结构分为2层, 芦苇(Phragmites communis)植株高1.0 ~ 2.0 m, 伴生种主要有香蒲(Acorus calamus)和藨草(Triglochin maritimum)。
16) 华扁穗草+西藏早熟禾+海韭菜群丛, 样地号7。为拉萨河谷典型的湿地草甸植被群落, 华扁穗草(Blysmus sinocompressus)为其建群种, 盖度70% ~ 80%, 草高10 cm左右, 无层次分化, 常伴生有西藏早熟禾(Poa tibetica)、海韭菜(Triglochin maritimum)、苔草等。
3.3 植被类型的分布与环境的关系图 2为拉萨半干旱河谷研究区域所有植物重要值的35个样地DCA二维排序结果, 沿对角线首先将不同生境的阴坡、阳坡分开, 不同生境的群落类型都有各自范围和分界线。各样地在坐标轴上的平面位置反映了样地所在群落的生态学特性(张金屯, 2004)。DCA所得的植物群落二维排序表明它们对植被与气候之间的关系分析较好。

|
图 2 拉萨半干旱河谷研究区域所有植物重要值的35个样地DCA二维排序 Figure 2 DCA ordination for all plant importance value of 35 plots of semi-arid valley region of Lasa Ⅰ:阔叶林Broadleaf forest, Ⅱ:灌丛Shrub, Ⅲ:草甸Meadow, Ⅳ:沼泽Swamp. |
DCA第1轴基本上反映各植物群落所在环境的海拔梯度变化趋势, 即从左到右, 海拔逐渐升高, 坡位向上, 由于气候差异、太阳辐射强弱不同、土壤类型不同以及降水、光照、湿度的相应变化, 在河谷研究区域植被随海拔的变化存在明显的分带; 海拔从底到高, 在河谷地带大致依次沼泽、落叶阔叶林、灌丛和草甸。
DCA第2排序轴反映了植物群落所处生境的水分状况, 表现出植物群落所在环境水分变化趋势, 即从下至上, 大气中水汽含量增加, 土壤中的水分含量逐渐增加, 即使在同一植被型中, 不同群丛所处生境的水分状况也不一样, 例如大果圆柏生长在水分条件较差的阳坡, 小半灌木草原、常绿革叶灌丛生境比落叶阔叶灌丛生境的水分优越; 到达一定的海拔高度(4 800 m以上), 阴坡和阳坡的生境接近, 植被类型相近。
根据拉萨半干旱河谷植物群落与环境因子排序, 不难看出, 水分和海拔是所有因子中对本山区植物群落分布起决定性作用的环境因子; 其次是坡向和坡位; 坡向对群落的分布也有较大关系。同时, DCA排序图较好地反映出山体各植物群落所在生境的环境状况。排序图与TWINSPAN分类结果基本吻合, 也验证了TWINSPAN分类结果的可靠性。
3.4 植被类型的垂直分布拉萨半干旱河谷主要植被类型的垂直分布见图 3。

|
图 3 拉萨半干旱河谷地带植被群落的垂直分布 Figure 3 Vertical distribution of semi-arid valley region communities of Lasa |
拉萨半干旱河谷研究区域植物群落的数量分类结果表明, 河谷地带由海拔、河流、坡向在温度、水分条件及两者的配合状况上存在着较大差别, 决定了半干旱河谷地带不同植物群落的分布类型与物种组成也不尽相同。采用生态重要值作为划分群落类型的依据(表 2, 3)。在半干旱河谷研究区域的1 350 m范围内垂直带谱上, 包括7个植被类型, 垂直带谱分明, 过渡带狭窄。海拔3 700 m左右是落叶阔叶林和沼泽植被群落, 海拔3 800 ~ 4 800 m阴、阳坡出现了不同的植物成分和群落类型。随着海拔的升高, 阴、阳坡的差异缩小, 植被群落类型和植物组成也相似。
拉萨半干旱河谷研究区域不同的生境和整个研究区域群落排序可以得到一致的结果, 这一点充分表明这一区域不同海拔各层次的物种组成都有其独特性, 各海拔的灌丛和草本层群落其特点明显, 每一类型的群落, 都具有其独特的“植物家族”, 群落植物种构成, 在较大程度上反映群落的物种构成特征。拉萨半干旱河谷植被分布, 由于受青藏高原独特的地形、地貌和气候影响, 植被分布规律与纬度地带性、经度地带性和垂直梯度相关, 环境胁迫强烈。
4 结论1) 应用TWINSPAN和DCA的分析方法, 对拉萨半干旱河谷地带植被群落进行研究, 得到与客观实际比较一致的分类结果。调查样方包括拉萨半干旱河谷群落植被类型, 共获35个样地。TWINSPAN分析结果表明, 研究区域的35个样地分为4个植被型组、7个植被型、14个群系、16个群丛, 研究区域植被类型多样, 垂直带分明。
2) 样地DCA第1排序轴反映了植物群落所在环境的海拔梯度, 在拉萨河谷植被随海拔的变化存在明显的分带, 过渡带狭窄。DCA第2排序轴反映了植物群落所处生境的水分状况, 从下至上, 土壤中的水分含量逐渐增加。对角线上各样地的排列顺序综合反映坡向、水分、热量、土壤等环境条件的差异。各群落类型都有自己的范围和分界线, 说明DCA排序较好的反映群落之间以及群落和环境之间的生态关系。
3) DCA排序结果与TWINSPAN分类结果具有一致性, 验证了TWINSPAN分类结果的准确性, 反映拉萨半干旱河谷地带主要植被分布是由海拔梯度、水分梯度和热量梯度等主要因子综合决定。
4) 以样地中植物的生态重要值为指标, 同时运用双向指示种分析和除趋势对应分析对植被进行定量分类, 能够更加客观地反映拉萨半干旱河谷区域的植被类型, 更好地了解该河谷植物群落的多样性特点, 为该地区地带性植被的恢复提供理论依据。
陈灵芝. 1993. 中国的生物多样性:现状及其保护对策[M]. 北京: 科学出版社.
|
代力民, 唐立娜, 曹玉明, 等. 2008. 辽东山区生态土地分类中的植物群落数量分析[J]. 林业科学, 44(3): 6-12. DOI:10.11707/j.1001-7488.20080306 |
郭东罡, 上官铁梁. 2005. 中条山中段植物群落数量分类与排序研究[J]. 武汉植物学研究, 23(5): 444-448. |
胡理乐, 毛志宏, 朱教君, 等. 2005. 辽东山区天然次生林的数量分类[J]. 生态学报, 25(11): 2848-2854. DOI:10.3321/j.issn:1000-0933.2005.11.008 |
江洪. 1994. 川西北甘南云杉林的DCA排序、环境解释和地理分布的模型研究[J]. 植物生态学报, 18(3): 209-218. |
马世骏. 1990. 现代生态学透视[M]. 北京: 科学出版社: 300-316.
|
青藏高原综合科学考察队. 1985. 西藏森林[M]. 北京: 科学出版社: 6-10.
|
青藏高原综合科学考察队. 1988. 西藏植被[M]. 北京: 科学出版社: 90-309.
|
席跃翔, 张金屯, 李军玲. 2004. 关帝山亚高山灌丛草甸群落的数量分类和排序研究[J]. 草业科学, 13(1): 15-20. |
熊咏梅, 朱纯, 何仲坚. 2008. 我国植物群落数量分类的研究进展[J]. 广东园林, (4): 49-51. |
杨继平. 2000. 世纪之交关注森林—林业的地位和作用[M]. 北京: 中国林业出版社: 88-97.
|
章家恩, 徐琪. 1997. 现代生态学研究的几大热点问题透视[J]. 地理科学进展, 16(3): 29-37. DOI:10.11820/dlkxjz.1997.03.005 |
章家恩, 徐琪. 1999. 恢复生态学研究的一些基本问题探讨[J]. 应用生态学报, 10(1): 109-113. |
周国逸. 1998. 关于恢复生态学[J]. 世界科技研究与发展, 20(1): 52-54. |
张金屯. 1995. 植被数量生态学方法[M]. 北京: 中国科学技术出版社.
|
张金屯. 2004. 数量生态学[M]. 北京: 科学出版社.
|
张峰, 张金屯. 2000. 我国植被数量分类和排序研究进展[J]. 山西大学学报:自然科学版, 23(3): 278-282. |
祝廷成, 钟章成. 1988. 植物生态学[M]. 北京: 高等教育出版社: 99-120.
|
Hill M O. 1979. TWINSPAN:A FORTRAN program for arranging multivariable data in an ordered two way table classification of the individuals and attributes ithaca[M]. N Y: Cornell University.
|
Lomolino M V. 2001. Elevation gradients of species-density historical and prospective views[J]. Global Ecology & Biogeography, 10(1): 3-13. |
ter Braak C J F.1988.CANOCO:A FORTRAN program for canonical community ordination by [partial][canonical ] correspondence analysis, principal analysis and redundancy analysis(Version2.1).Wageningen:Agricultural Mathematics Group, Ministry of Agriculture and Fisheries. http://www.numdam.org/item?id=MSH_1999__147__135_0
|
Zdravko B. 2005. Vegetation-environment relationships and classification of the seasonal savannas in Venezuela[J]. Flora, 200(1): 49-64. DOI:10.1016/j.flora.2004.06.001 |