 2010, Vol. 46
2010, Vol. 46文章信息
- 周亮, 刘盛全, 刘亚梅, 刘倩
- Zhou Liang, Liu Shengquan, Liu Yamei, Liu Qian
- 欧美杨107杨生长应变与木材解剖特征和干缩性质之间的关系
- Relationships between Growth Strain and Anatomical Characters and Shrinkage Properties in Poplar Clone 107(Populus × euramericana cv.'Neva') Tree
- 林业科学, 2010, 46(9): 124-129.
- Scientia Silvae Sinicae, 2010, 46(9): 124-129.
-
文章历史
- 收稿日期:2009-01-06
- 修回日期:2009-07-27
-
作者相关文章



生长应力产生于树木次生生长过程中,主要是树木为了保持其主干和树枝在原有状态下生长而形成了应力保护机制(Archer,1986;Chafe,1979)。它的存在对木材加工利用带来了一定的影响,主要是由于高生长应力和生长应力分布不均导致原木易发生端裂,制材过程中板材出现开裂、横弯及扭曲等缺陷,从木材利用角度,严重制约了锯材的出材率和高附加值实木的利用(Yang et al., 2001)。为了研究生长应力对木材加工利用的影响,学者们一方面研究生长应力的大小和分布与制材缺陷之间的关系(Jullien et al., 2003;Okuyama et al., 2004),另一方面着手研究生长应力与木材性质之间关系(Clair et al., 2003b;Malan,1987;Malan et al., 1987)。通过这种关系的研究不仅可以分析生长应力大小对木材性质的影响,还可以通过它们之间的相互关系对生长应力或木材性质进行预测。Malan等(1987)通过对正常生长巨桉(Eucalyptus grandis)研究认为木材性质与生长应变水平之间没有显著的相关性;胡继青等(2000)对3种正常生长桉树[巨尾桉(E.grandis×E.urophylla),尾园桉(E.urophylla×E.tereticornis),粗皮桉(E.pellita)]研究也发现生长应变与基本密度、生材密度无关;刘晓丽(2005)在正常生长尾巨桉(E.urophylla×E.grandis)研究中得出,生长应变与轴向干缩率有显著正相关关系,而与基本密度和MOE等性质之间关系较弱;Fang等(2008)通过对不同倾斜角度下的Ⅰ-69杨(Populus deltoids cv.“Lux”ex Ⅰ-69/55)的研究发现,生长应力指示值与大多数木材材质指标之间关系显著;Clair等(2003b)在欧洲板栗(Castanea sativa)研究中,分别对含有胶质层纤维的应拉木和正常木的生长应力指示值与木材物理力学性质之间关系展开研究,发现了应拉木与正常木所得到的结论存在较大的不同。
欧美杨107杨(Populus×euramericana cv.‘Neva’)目前在我国华北和黄淮等区域广为栽植,已经成为我国人工林中较为重要的杨树优良品系之一(张绮纹等,2003)。前文以欧美杨107杨为研究材料,针对正常生长和偏心生长2种不同条件下的生长应变分布情况展开了论述(周亮等,2009)。在此基础上,本文将讨论2种生长条件下立木生长应变与木材性质之间的关系,以期为欧美杨107杨加工利用提供必要的理论信息。
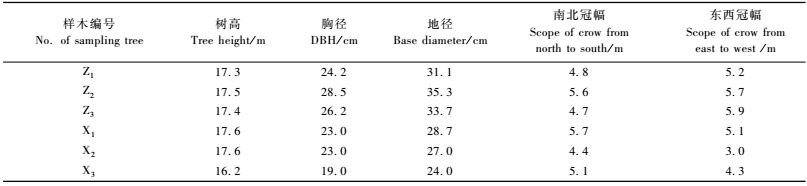
1 材料与方法 1.1 试材采集采集地位于河南省濮阳市白岗乡林场,该林场位于黄河滩区,为黄河冲积平原,土壤类型为轻沙壤,偏盐碱,pH 7.8左右。按照小班数据进行林地踏查,挑选3株长势良好、干形较为通直的7年生欧美杨107杨平均木,依次编号为Z1,Z2,Z3;从林地东西向主干渠两旁选择了3株具有不同倾斜角度或弯曲程度的7年生欧美杨107杨,依次编号为X1,X2,X3。样木生长情况如表 1所示。
|
|
选取Z1,Z2 2个单株的1.5 m处和Z3,X1,X2、X3 4个单株0.5,1.5.2.5 m 3个不同高度处进行立木表面生长应变测定,在树干周向上具体测试位置的选择参见周亮等(2009)。采用应变片法对相应位置进行立木表面轴向生长应变的测定(Yoshida et al., 2002)
1.2.2 木材纤维形态测定将应变测试点下方试块从立木中取出,采用离析法测定树干最外围纤维长度(每个试样共测定50根纤维的长度,取平均值),并将试块制作常规木材切片,测定纤维宽度、双壁厚,并计算胞腔径、长宽比、壁腔比。
1.2.3 木材干缩性质测定将测试点下木材制备成尺寸大小为1 cm×1 cm×2 cm的小试块,然后按照国家标准(GB 1932-1991)测定木材的轴向全干干缩率、径向全干干缩率、弦向全干干缩率。
1.2.4 数据分析方法采用SPSS软件对数据进行统计分析。
2 结果与分析 2.1 正常生长立木表面轴向生长应变与木材性质之间的关系将正常生长立木各测试点的生长应变值与相对应试块测定的纤维形态指标和干缩性指标作散点图,并添加线性趋势线,结果如图 1所示。随着生长应变数值的增加,纤维长度、胞腔径、长宽比、径向全干干缩率、弦向全干干缩率出现微弱的下降趋势,而纤维宽度、双壁厚、壁腔比、轴向全干干缩率同时出现微弱的上升趋势。反映到生长应力上,则表现为:随着生长拉应力的增加,纤维长度、胞腔径、长宽比、径向全干干缩率、弦向全干干缩率减小;纤维宽度、双壁厚、壁腔比、轴向全干干缩率增加。计算各测试点生长应变与纤维形态指标和干缩性指标之间的相关系数,结果如表 2所示。生长应变与所有纤维形态指标和干缩性指标之间的相关系数均不显著。此外,反映出来的相关系数方向与散点图得出的结果一样。

|
图 1 正常生长立木表面生长应变与木材性质之间关系 Figure 1 Correlation between surface longitudinal growth strain and wood properties of normal growth tree |
|
|

从本文研究结果来看,生长应变与所测定木材性质指标之间关系均不显著,且相关系数也较小,这说明了正常生长欧美杨107杨生长应变与木材纤维形态和干缩性质之间的关系较为微弱。这一结果与前人的相关结果基本一致。纤维形态与应变之间的关系研究结论与Malan等(1987)对桉树研究较为一致,而在干缩性质研究与刘晓丽(2005)对桉树的研究存在不同,本文中各干缩指标与生长应变之间关系均不显著,这一结论和Clair等(2003b)对直立生长的欧洲栗研究结果一致。
2.2 偏心生长立木生长应变与相关木材材性之间的关系将偏心生长立木各测试点的生长应变值与相对应试块测定的纤维形态指标和干缩性指标作散点图,并添加线性趋势线,结果如图 2所示。随着生长应变数值的增加,双壁厚、长宽比、径向全干干缩率、弦向全干干缩率、轴向全干干缩率出现较快的下降趋势,而纤维宽度、胞腔径同时出现较快的上升趋势,纤维长度出现微弱的上升趋势。反映到生长应力上,则表现为:随着生长拉应力的增加,双壁厚、长宽比、径向全干干缩率、弦向全干干缩率、轴向全干干缩率迅速减小;纤维宽度、胞腔径迅速增加;纤维长度缓慢增加。计算各测试点生长应变与纤维形态指标和干缩性指标之间的相关系数,结果如表 2所示。偏心生长立木表面轴向生长应变与所有纤维形态指标和干缩性指标之间的相关系数结果显示:生长应变与双壁厚、轴向全干干缩率、径向全干干缩率、弦向全干干缩率、壁腔比的负相关关系在0.001水平上显著,与胞腔径的正相关关系在0.001水平上显著,与纤维宽度的正相关关系在0.01水平,与弦向全干干缩率的负相关关系在0.01水平上显著,与长宽比的负相关关系在0.05水平上显著,与纤维长度之间相关系数不显著。

|
图 2 偏心生长立木表面生长应变与木材性质之间关系 Figure 2 Correlation between surface longitudinal growth strain and wood properties of eccentric growth tree |
从本文研究结果来看,偏心生长欧美杨107杨立木生长应变与多数木材纤维形态和干缩性质指标之间关系较为紧密,尤其是双壁厚、轴向全干干缩率、径向全干干缩率、弦向全干干缩率、壁腔比、胞腔径等材性指标。Fang等(2008)在对Ⅰ-69杨研究中得出的生长应力指示值与木材性质之间关系结果与本文所得的研究结果基本一致,主要差别在于弦向全干干缩、径向全干干缩和纤维长度的研究结果上。本文中弦向干缩和径向干缩与应变之间为显著正相关,而Fang在对Ⅰ-69杨研究中得出生长应力指示值与弦向干缩和径向干缩显著负相关;Clair等(2003b)对偏心生长欧洲栗研究中得出生长应力指示值与弦向干缩为显著正相关,与本文结论相同。本文研究认为偏心生长树干最外层纤维长度与测定的应变之间关系不显著,Fang研究得出纤维长度与生长应力指示值为显著正相关。上述差异性可能来源于诸多种因素之间的差异,如研究材料、测试方法、木材材性测试取材部位等。此外,关于应拉木和正常木纤维长度之间的比较结果,不同的学者由于研究材料和取材方法之间的差异,得出的结论也不经相同(Jourez et al., 2001;Zimmermann 1964)。
学者们普遍认为多数偏心生长的立木可能在年轮较宽区域出现应力木,而应力木的存在对生长应力的影响也得到了诸多证实(Du et al., 2007;Wardrop,1956;Wardrop et al., 1948)。一般认为,在阔叶材应拉木区域表现为拉应力较大,在针叶材应压木区域表现为拉应力较小或者直接表现为较大的压应力(Bamber 2001;Yamamoto et al., 2002)。因此学者们在研究生长应力与木材性质之间关系时,对研究材料的选择也大致分成两大类:一类是正常生长的木材(通常为直立生长的树干),另一类是偏心生长的木材(通常树干中出现倾斜或者弯曲)。在第2类材料中通常会有一定量的应力木存在,而第1类材料中应力木含量很少甚至不存在。由于应力木与正常木材质之间存在较大的差异,如应拉木中由于胶质层的存在而形成的细胞壁较厚、腔小(Wardrop,1956;Wardrop et al., 1948),以及S1层在细胞壁中组成较大而引起的木材干缩性质变化等(Clair et al., 2003a;Fang et al., 2007),因此在分析偏心生长木材材性与生长应力之间关系时,往往是生长应力水平与应力木存在程度之间相互交织,共同影响了木材材质。
综合比较本文对正常生长立木和偏心生长立木2种不同材料得出的研究结果,可以发现两者之间的差异较大。正常生长的立木其本身在不同树干位置和不同单株之间的应变数值差异较小,此外,在没有应力木干扰的情况下,其树干外围材性的变异也有限,这些因素可能是导致生长应力与木材材性指标之间关系不明显的原因之一。然而,偏心生长立木其本身应变数值在不同单株和不同位置之间差异较大,而这种差异的变化规律与应拉木存在程度又紧密相连,再加上应拉木与正常木在木材材性指标之间的差异,必然导致了生长应变与多数纤维形态指标和木材干缩性质指标之间关系明显。因此,本文认为对于不含应拉木正常生长的立木,其生长应力与木材纤维形态和木材干缩性质之间关系较为微弱;对于含应拉木偏心生长的立木,由于应拉木材性与正常木之间的差别,其生长应力与与多数木材纤维形态指标和木材干缩性质指标之间关系较为紧密。
3 结论随着生长应变数值的增加,正常生长立木纤维长度、胞腔径、长宽比、径向全干干缩率、弦向全干干缩率出现微弱的下降趋势,而纤维宽度、双壁厚、壁腔比、轴向全干干缩率同时出现微弱的上升趋势。正常生长立木生长应变与所有纤维形态指标和干缩性指标之间的相关系数均不显著。
随着生长应变数值的增加,偏心生长立木双壁厚、长宽比、径向全干干缩率、弦向全干干缩率、轴向全干干缩率出现呈现较快的下降趋势,而纤维宽度、胞腔径同时出现较快的上升趋势,纤维长度出现微弱的上升趋势。偏心生长立木生长应变与双壁厚、轴向全干干缩率、径向全干干缩率、弦向全干干缩率、壁腔比的负相关关系在0.001水平上显著,与胞腔径的正相关关系在0.001水平上显著,与纤维宽度的正相关关系在0.01水平,与弦向全干干缩率的负相关关系在0.01水平上显著,与长宽比的负相关关系在0.05水平上显著,与纤维长度之间相关系数不显著。
对于不含应拉木正常生长的立木,其生长应力与木材纤维形态和木材干缩性质之间关系较为微弱;对于含应拉木偏心生长的立木,由于应拉木材性与正常木之间的差别,其生长应力与多数木材纤维形态指标和木材干缩性质指标之间关系较为紧密。
胡继青, 姜笑梅, 侯祝强, 等. 2000. 三种人工林桉树轴向生长应变变异初探[J]. 木材工业, 14(6): 9-11. |
刘晓丽. 2005. 人工林尾巨桉生长应变与木材性质关系及高生长应变形成机理的研究. 中国林业科学研究院博士学位论文.
|
张绮纹, 李金花. 2003. 杨树工业用材林新品种[M]. 北京: 中国林业出版社.
|
周亮, 刘盛全, 刘倩, 等. 2009. 欧美杨107杨立木生长应变分布规律[J]. 林业科学, 45(1): 118-124. DOI:10.11707/j.1001-7488.20090121 |
Archer R R. 1986. Growth stresses and strains in trees. Springer Verlag, Berlin.
|
Bamber R K. 2001. A general theory for the origin of growth stresses in reaction wood: how trees stay upright[J]. IAWA Journal, 22(3): 205-212. DOI:10.1163/22941932-90000279 |
Chafe S C. 1979. Growth stress in trees[J]. Australian Forest Research, 9(3): 203-223. |
Clair B, Jaouen G, Beauchêne J, et al. 2003a. Mapping radial, tangential and longitudinal shrinkages and relation to tension wood in discs of the tropical tree Symphonia globulifera[J]. Holzforschung, 57(6): 665-671. |
Clair B, Ruelle J, Thibaut B. 2003b. Relationship between growth stresses, mechano-physical properties and proportion of fibres with gelatinous layer in chestnut (Castanea Sativa Mill.)[J]. Holzforschung, 57(2): 189-195. |
Du S, Yamamoto F. 2007. An overview of the biology of reaction wood formation[J]. Journal of Integrative Plant Biology, 49(2): 131-143. DOI:10.1111/jipb.2007.49.issue-2 |
Fang C H, Clair B, Gril J, et al. 2007. Transverse shrinkage in G-fibers as a function of cell wall layering and growth strain[J]. Wood Science and Technology, 41(8): 659-671. DOI:10.1007/s00226-007-0148-3 |
Fang C H, Guibal D, Bruno C, et al. 2008. Relationship between growth stress and wood properties in poplar Ⅰ-69 (Populus deltoides Bartr. cv."Lux"ex Ⅰ-69/55)[J]. Annals of Forest Science, 65(3): 1-9. DOI:10.1051/forest:2008999 |
Jullien D, Laghdir A, Gril J. 2003. Modelling log-end cracks due to growth stresses: calculation of the elastic energy release rate[J]. Holzforschung, 57(4): 407-414. |
Jourez B, Riboux A, Leclercq A. 2001. Anatomical characteristics of tension wood and opposite wood in young inclined stems of poplar (Populus euramericana cv. Ghoy)[J]. IAWA Journal, 22(2): 133-157. DOI:10.1163/22941932-90000274 |
Malan F S. 1987. The reduction of high levels of growth stress in Eucalyptus grandis-15 tree breeding our only solution[J]. Wood Southern Africa, 12(1): 19-20. |
Malan F S, Gerischer G F R. 1987. Wood property differences in South African grown Eucalyptus grandis trees of different growth stress levels[J]. Holzforschung, 41(6): 331-335. DOI:10.1515/hfsg.1987.41.6.331 |
Okuyama T, Doldan J, Yamamoto H, et al. 2004. Heart splitting at crosscutting of eucalypt logs[J]. Journal of Wood Science, 50(1): 1-6. DOI:10.1007/s10086-003-0533-y |
Wardrop A B. 1956. The nature of reaction wood. V. The distribution and formation of tension wood in some species of Eucalyptus[J]. The distribution and formation of tension wood in some species of Eucalyptus. Australian Journal of Botany, 4(2): 152-166. |
Wardrop A B, Dadswell H E. 1948. The nature of reaction wood Ⅰ—The structure and properties of tension wood fibres[J]. Aust J Sci Res B: 1-16. |
Yamamoto H, Yoshida M, Okuyama T. 2002. Growth stress controls negative gravitropism in woody plant stems[J]. Planta, 216(2): 280-292. DOI:10.1007/s00425-002-0846-x |
Yang J L, Waugh G. 2001. Growth stress, its measurement and effects[J]. Australian Forestry, 64(2): 127-135. DOI:10.1080/00049158.2001.10676176 |
Yoshida M, Okuyama T. 2002. Techniques for measuring growth stress on the xylem surface using strain and dial gauges[J]. Holzforschung, 56(5): 461-467. |
Zimmermann M H. 1964. The formation of wood in forest trees[M]. Academic Press, New Yor.
|